
Translate this page into:
Effect of aluminium on lipid profile and atherogenic index in prepubertal and young adult female rats: A pilot study
*Corresponding author: Prasunpriya Nayak, Department of Physiology, All India Institute of Medical Sciences, Jodhpur, Rajasthan, India. nprasunpriya@gmail.com
-
Received: ,
Accepted: ,
How to cite this article: Ghosh S, Gaur A, Sengupta T, Banerjee M, Nayak P. Effect of aluminium on lipid profile and atherogenic index in prepubertal and young adult female rats: A pilot study. Indian J Physiol Pharmacol 2023;67:92-9.
Abstract
Objectives:
Exposure of aluminium (Al) to mankind is inescapable, and its dyslipidaemic impact is a possible contributing factor to health hazards like cardiovascular diseases. The health effects due to the metalloestrogenic property of the metal need imperative consideration. The current experimental work was undertaken to explore Al-induced dyslipidaemia due to its metalloestrogenic property.
Materials and Methods:
To fulfil this objective, prepubertal (PP) and young adult (YA) female Wistar rats were intraperitoneally administered to two doses of Al [5 and 10 mg/Kg body weight (BW)] once daily for 2 weeks. After the completion of the acute exposure protocol, plasma and hepatic tissue lipid profiles were estimated. Analysis of variance was carried out by the Kruskal–Wallis test and the differences between the groups were analysed by Mann–Whitney U post hoc test
Results:
Increased triglyceride, total cholesterol, low-density lipoprotein (LDL) cholesterol and very-LDL cholesterol in plasma were found in YAs treated with both doses of Al in a dose-dependent manner. Similar changes were not present in PP female rats. Decreased levels of lipid levels were observed in the case of hepatic lipid profile.
Conclusion:
The study ushers light towards the dyslipidaemic alterations in experimental female rats after acute Al exposure. Impacts of Al on the growth and organosomatic index during the vital developmental days were significantly decided by the pubertal status of the female rats. The results of this study indicate the impact of puberty on the Al-induced modifications in lipid profile parameters and cardiovascular risk factors.
Keywords
Aluminium
Dyslipidaemia
Oestrogen
Liver
Atherogenic index
Puberty

INTRODUCTION
Aluminium (Al) is an extensively scattered metal in the environment,[1] naturally occurring in its trivalent state (Al+3) forming organic and inorganic complexes.[2,3] Even though it is an abundantly present metal, the useful physiological role of Al in the human body is not reported, and its exposure is unsafe.[4] Excess Al exposure causes toxic effect which perturbs the entire physiology, including alteration of metabolic pathways, like lipid metabolism.[5] Considering the current worldwide health scenario,[6] dyslipidaemia and the associated health problems need urgent attention.
Dyslipidaemia is linked with various neurological and cardiovascular diseases.[7,8] Interestingly, gender has a role in the lipid profile of dyslipidaemic patients.[9] Before menopause, females have a lower risk of atherosclerotic cardiac disease compared with the age-matched males. Reports indicate that women have a 10-year delay in the first event of myocardial infarction compared to men.[10,11] Unlike males, different dyslipidaemic indices include reduced low and very-low-density lipoproteins (LDL and VLDL, atherogenic factor) and raised high-density lipoproteins (HDL, atheroprotective factor) in women contribute to a lower risk of cardiovascular disorder in them.[12]
The steroid hormones, mainly oestrogen have been proposed to contribute to these differences in lipid levels and metabolism in and around puberty.[13-16] There is also evidence that in women the time of puberty onset leads to altered lipid levels.[17] Following menopause, a reduction in circulating oestrogen levels increases susceptibility to developing metabolic disorders in women.[16] Exposure to many environmental and synthetic products (endocrine disruptors) during and before puberty affects pubertal timing and leads to long-term disturbance to the hypothalamo– pituitary–gonadal axis.[18,19] Oestrogen-like endocrine disruptors cause disturbed metabolic effects by intruding with the synthesis, metabolism and/or binding of natural oestrogens.[20-22]
Metal ions meddling with oestrogen action have been termed metalloestrogen (ME), some of whose physiological roles are implicit.[23] MEs enhance the proliferation of cells that are oestrogen-dependent for growth.[24] The relationship between in utero and peripubertal metal exposure with reproductive hormone levels and sexual progression among adolescent girls in Mexico has been investigated. It was found that various metals, including Al, were associated with delayed pubertal progression in the exposed girls.[25] Moreover, Al has been shown to differentially affect anxiety-like behaviour in rats of two different age groups when exposed during and before puberty.[26] The possible association between Al as a leading cause of dyslipidaemia has also been indicated.[27] However, there is a paucity of literature explaining whether and how an environmental toxin like Al, causes dyslipidaemia in and around puberty. Thus, the present study was conceived to evaluate the Al-induced changes in lipid profile in female rats before and after puberty.
MATERIALS AND METHODS
Animals
The experiment was conducted on 18 prepubertal (PP) female Wistar rats (age 30–35 days), and 18 young adults (YAs) female Wistar rats (age 55–60 days). The rats were obtained from the Animal House facility of the Department of Zoology, Jai Narayan Vyas University, Jodhpur. The experimental protocol was approved by the Institutional Animal Ethics Committee (1646/CO/ReBi/8/12/CPCSEA), on 15.9.2020. All the procedures were performed according to the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals, India. Just after weaning (approximately 25 days from birth), the PP female rats were recruited for the study, the YA female rats were 50 days from birth.
Methodology
Aluminium chloride was procured from Sigma, India. Other chemicals were of analytical grade. The Al-0 group served as control and was treated with normal saline (i.p.), and the Al-5 and Al-ten groups were exposed to Aluminium chloride (AlCl3) solution (intraperitoneal i.p.), at doses of 5 and 10 mg Al/kg body weight (BW), respectively. The low dose of 5 mg/ kg BW dose of AlCl3 solution exposures was selected from the previous studies[26,28] and the other dose was double that of the already reported dose. Both age groups of rats were divided with the help of Random Allocation Software (Version 1.0, May 2004) into three subgroups (Al-0, Al-5 and Al-10), having six animals in each. All the six groups of rats, Al-0, Al-5 and Al-10 groups of PP and YA females, were administered with either saline or AlCl3 at the two doses for 2 weeks at the same time of the day. The rats were provided with standard laboratory rat chow and drinking water ad libitum throughout the study. The detailed study design is presented in [Figure 1].
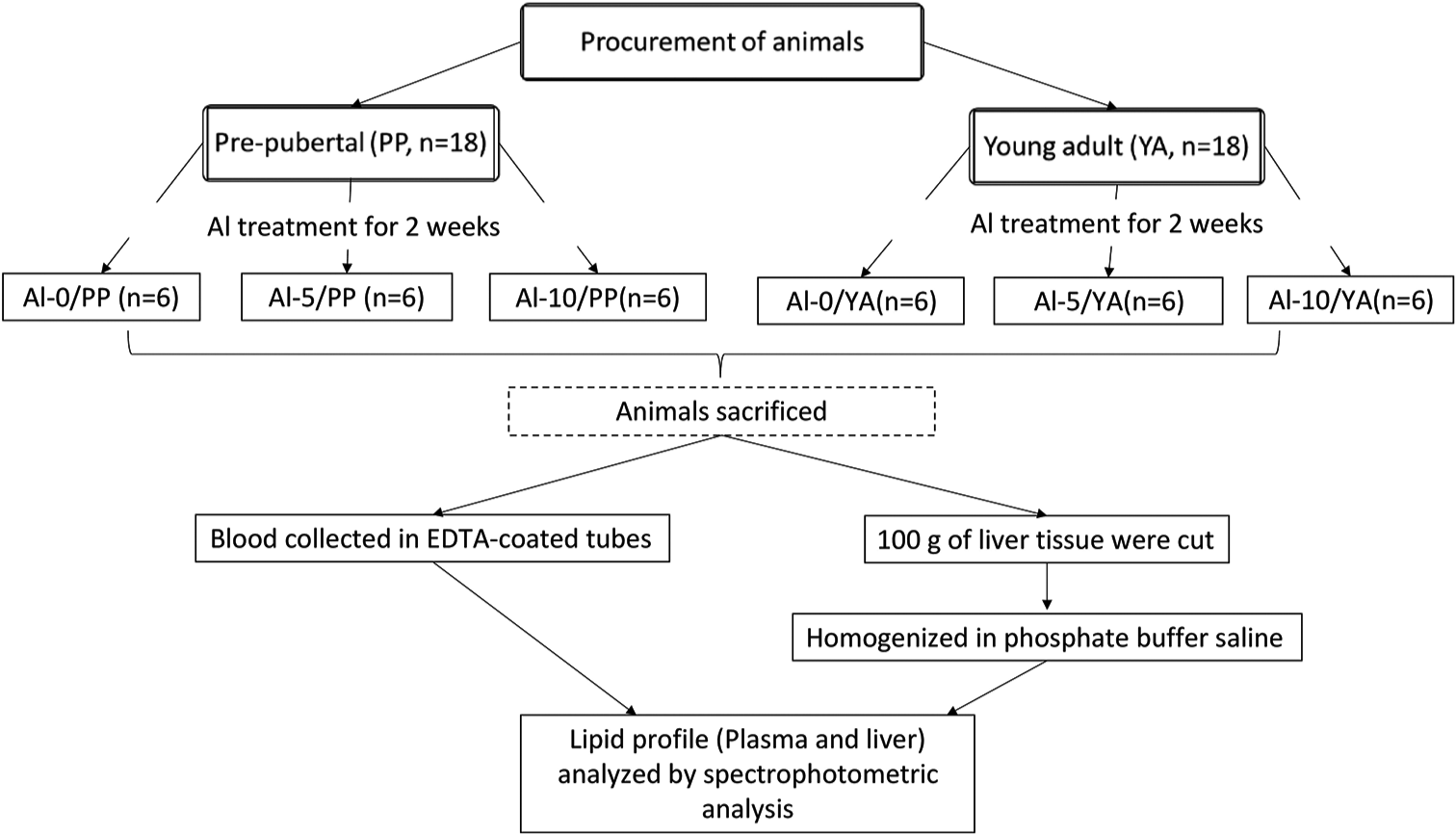
- The experimental design. PP: Prepubertal, YA: Young adult, Al-0: Control, Al-5: 5 mg/kg Aluminium, Al-10: 10 mg/kg Aluminium, EDTA:Etheylene diamine tetra-acetic acid, n: Number.
After 2 weeks of treatment, blood samples from overnight fasted rats were collected through the retro-orbital puncture and the rats were sacrificed by cervical dislocation. Blood was collected in ethylenediaminetetraacetic acid-coated tubes and centrifuged at 3000 revolutions per minute (RPM) for 15 min at room temperature. The plasma obtained was used to determine the fasting lipid profile. The blood sample for lipid profile was analysed by spectrophotometric analysis (Beckman Coulter analyser, CA, USA) by cholesterol oxidaseperoxidase method (Agappe Diagnostic Ltd, New Delhi). The same enzymatic analysis along with immunoinhibition assay was employed to estimate HDL (Agappe Diagnostic Ltd, New Delhi) and TG (Agappe Diagnostic Ltd, New Delhi). All the protocol was followed as per the manufacturer’s instruction. Serum LDL and VLDL levels were calculated as per the formula described previously.[29]
LDL = Total Cholesterol - ([TG]/5+HDL)
VLDL = TG/5
After weighing the whole liver, approximately 100 g of liver tissue was cut, minced and homogenised in phosphate buffer saline (pH 7.4) using the Teflon homogeniser. The total lipid profiling from the liver sample was also done by spectrophotometric kit following the manufacturer’s protocol as adapted from Folch et al.,[30] Plasma and liver samples were preserved at −40°C. Biochemical processing of plasma and liver homogenates for lipid profile was carried out within 2 days of sacrifice.
From the estimated biochemical parameters, the atherogenic indices (AI) were also calculated, which are – Castelli risk index-I (CRI-I = TC/HDL-C), Castelli Risk Index-II (CRI-II = LDL-C/ HDL-C), atherogenic coefficient (AC = [TC–HDL-C]/HDL-C) and atherogenic index of plasma (AIP = log [(TG)/HDL-C]).[31]
Statistical analysis of data
Analysis of variance was carried out by Kruskal–Wallis (KW) test and the differences between the groups were analysed by the Mann–Whitney U post hoc test accepting the probability of 5% or less as significant using PAST statistical software (ver. 4.03; Copyright: Ø. Hammer 1999–2020).[32] To present the data graphically, box and whisker plots have been used. The median value (bold horizontal line), interquartile range (boxes on either side of the line) and the outliers (small circles) if any, are also indicated.
RESULTS
Two age groups of female Wistar rats (PP and YA) were exposed to two different doses of AlCl3 and maintained along with the same age groups of control rats. Although, no mortality was recorded during the experimental period; Al-10 treated rats were relatively less active compared to animals of the other two groups (Al-0 and Al-5). No difference in group-wise food and water consumption was noted (data not shown). The Kruskal-Wallis (KW) test detected a significant difference (P = 0.01) between the BW gain of PP rats (Al-0, Al-5 and Al-10) but not in YA groups (P = 0.1), as shown in [Table 1]. The percentage gain of BW during the experimental period was significantly higher in the Al-5-treated PP rats [Table 1]. Similarly, a higher organosomatic index of the liver was noted in the PP rats treated with Al-5 [Table 1] in comparison to the PP Al-0 group only (P = 0.01) and the PP groups showed a significant (P = 0.03) difference in the KW test.
| Animal groups | Al-0 group | Al-5 group | Al-10 group |
|---|---|---|---|
| Change in BW (g%) during the exposure period | |||
| Prepubertal | 33.52±4.67 | 64.83±3.56a | 32.74±6.44c |
| Young adult | 24.64±2.71b | 20.19±2.04b | 15.51±3.29b |
| Post-exposure liver weight (g/100g BW) | |||
| Prepubertal | 4.48±0.14 | 5.75±0.23a | 5.52±0.46a |
| Young adult | 5.30±0.29b | 4.59±0.15ab | 5.02±0.13 |
Data are presented as Mean±standard error of the mean (n=6). aindicates significant difference with the Al-0 subgroup of the same age group of rats. bindicates a significant difference with the prepubertal rats of the same Al exposure group. cindicates a significant difference with the Al-5 subgroup of the same age group of rats. BW: Body weight, Al: Aluminium, n: Number.
Comparison of serum lipid profiles
All the groups of YA female rats showed significantly higher levels of plasma TG compared to the PP female rats (χ2 = 28.7, P < 0.01) [Figure 2]. The Al-exposed YA female rats demonstrated a significantly higher level of plasma TG compared to their control counterpart (P = 0.01), and the plasma TG of Al-10 was significantly higher than the Al-5 YA rats (P = 0.02) [Figure 2a]. Significantly higher plasma cholesterol levels were seen in both Al-exposed YA rats (χ2 = 27.3, P < 0.01). Insignificant alterations in plasma cholesterol levels were observed in both doses of Al-exposed PP female rats [Figure 2b]. In the case of plasma VLDL cholesterol [Figure 2c], the alterations are very similar to that of plasma TG. Statistically significant higher level of plasma VLDL cholesterol in Al-exposed YA females compared to PP female rats in a dose-dependent manner is shown (χ2 = 28.7, P < 0.01) in [Figure 2c]. The plasma LDL cholesterol levels in control YA females were lower than that of PP control female rats (P = 0.01) [Figure 2d]. These plasma LDL cholesterol levels in control YA female rats were lower than the Al-exposed rats of the same age group; however, it was statistically significant compared to the Al-5 group (P = 0.01) but insignificant when compared to the Al-10 group.

- Box plot for plasma (a) triglyceride, (b) total cholesterol, (c) VLDL-Cholesterol, (d) LDL cholesterol and (e) HDL cholesterol contents of prepubertal (n: number, n=6 for Al-5 and Al-10 each) and young adult (n: number, n=6 for Al-5 and Al-10 each) female rats exposed to two doses of aluminium (Al). # indicates a significant difference with the prepubertal rats of the same Al exposure group. @ indicates a significant difference with the Al-0 subgroup of the same age group of rats. $ indicates a significant difference with the Al-5 subgroup of the same age group of rats. LDL: Low-density lipoprotein, VLDL: Very-low-density lipoprotein, HDL: High-density lipoprotein.
From [Figure 2], it is observed that the plasma lipid parameters were undisturbed by the Al exposure to PP female rats; however, plasma HDL cholesterol levels were significantly elevated in the Al-5 group (P = 0.01) and insignificant in the Al-10 group of PP female rats [Figure 2e]. A similar trend of raised plasma HDL cholesterol was observed in Al-exposed YA female rats, but the differences between the groups were statistically insignificant.
Comparison of hepatic lipid profiles
The TG levels in the liver of YA female rats were also significantly higher (χ2 = 31.6, P < 0.01) compared to their respective exposure group in PP female rats [Table 2]. There was a ten-fold rise in the liver TG levels of the control YA females compared to that of the Al-exposed rats. The liver TG levels were also significantly higher in Al-10 groups in comparison to the Al-5 groups of female rats (P = 0.4) irrespective of their reproductive age. Similar trends in liver total cholesterol (χ2 = 26.9, P < 0.01), VLDL cholesterol (χ2 = 31.6, P < 0.01) and HDL cholesterol (χ2 = 15.8, P = 0.004) were observed in response to the two doses of Al exposure in the two age groups of female rats [Table 2]. Compared to the control female rats, respectively, higher and lower levels of liver LDL cholesterol were observed in Al-10 groups of PP and YA female rats. However, there is no significant difference in the hepatic tissue LDL-C levels of the Al-10 group when compared with the other group of rats.
| Animal groups | Al-0 group | Al-5 group | Al-10 group |
|---|---|---|---|
| Total liver triglyceride (mg) | |||
| Prepubertal | 10.57±0.94 | 8.63±1.65 | 22.01±1.66ac |
| Young adult | 133.19±23.30b | 33.81±2.00ba | 53.69±7.63bac |
| Total liver cholesterol (mg) | |||
| Prepubertal | 2.11±0.18 | 1.72±0.33 | 4.40±0.33ac |
| Young adult | 26.63±4.66b | 6.76±0.40ba | 10.73±1.52bac |
| Total liver LDL cholesterol (mg) | |||
| Prepubertal | 0.52±0.09 | 0.43±0.09 | 1.39±0.39 |
| Young adult | 1.85±0.18b | 1.27±0.27b | 1.55±0.15 |
| Total liver HDL cholesterol (mg) | |||
| Prepubertal | 0.03±0.03 | 0.11±0.05a | 0.25±0.00ac |
| Young adult | 1.17±0.25b | 0.20±0.15a | 0.21±0.11a |
Data are presented as Mean±standard error of the mean (n=6). aindicates significant difference with the Al-0 subgroup of the same age group of rats. bindicates a significant difference with the prepubertal rats of the same Al exposure group. cindicates a significant difference with the Al-5 subgroup of the same age group of rats. LDL: Low-density lipoprotein, HDL: High-density lipoprotein, Al: Aluminium, n: Number
Comparison of lipid ratios and atherogenic indices
The lipid ratios and atherogenic indices such as Castelli’s Risk Index I (CRI-I), Castelli’s Risk Index II (CRI-II), AC and AI of plasma are presented in [Table 3]. When compared between the Al-exposed and control groups, no significant alterations were observed in the indices in either of the study groups. The Al-5 and Al-10 groups demonstrated significantly raised indices (CRI-I, χ2 = 16.8, P = 0.005) and (AIP, (χ2 = 24.5, P= 0.0001) in YA female rats compared to PP female rats.
| Animal groups | Al-0 group | Al-5 group | Al-10 group |
|---|---|---|---|
| Castelli’s risk index-I | |||
| Prepubertal | 1.76±0.03 | 1.82±0.06 | 1.91±0.12 |
| Young adult | 1.81±0.30 | 2.29±0.14b | 2.44±0.08b |
| Castelli’s risk index-II | |||
| Prepubertal | 0.49±0.04 | 0.47±0.05 | 0.53±0.03 |
| Young adult | 0.35±0.09 | 0.56±0.06 | 0.49±0.10 |
| Atherogenic coefficient | |||
| Prepubertal | 0.76±0.03 | 0.82±0.06 | 0.91±0.12 |
| Young adult | 0.81±0.30 | 1.29±0.14b | 1.44±0.08b |
| Atherogenic index | |||
| Prepubertal | 0.19±0.05 | 0.14±0.05 | 0.23±0.04 |
| Young adult | 0.59±0.09b | 0.58±0.04b | 0.68±0.07b |
Data are presented as Mean±standard error of the mean (n=6). bindicates a significant difference with the prepubertal rats of the same Al exposure. Al: Aluminium, n: Number
DISCUSSION
The exposure to Al caused a significant increase in BW in the Al-5 group of PP female rats. Contrary to this, no significant change in BW gain was observed in both the Al exposed groups compared to the control YA female rats. Wide variations were noted in these data which did not meet statistical significance. These data suggest that an acute Al exposure for just 2 weeks might affect the normal growth and metabolism. Further to corroborate the finding, and to appreciate the metalloestrogenic property of Al exposure, the two doses of Al exposure raised the organosomatic index of the liver in PP female rats only. There are inconsistent reports of Al-induced changes in BW where the exposure duration varied from 4 weeks to lifelong.[33,34] The acute exposure to Al was limited to 2 weeks as that was the window period available for the PP exposure group between weaning and adolescence.[35]
In humans, variable changes in fasting lipid profile during puberty have been documented earlier.[14] We have observed a post-pubertal increase in plasma TG and VLDL cholesterol levels in the YA female rats. The Al-treated YA female rats had higher plasma lipid parameters compared to the PP control group (Al-0/PP). All the liver lipid parameters were relatively lower in Al-exposed YA female rats in comparison to the age-matched control group (Al-0/YA). However, this was not the type of observation seen in PP female rats. The Al-10/PP female rats had higher liver lipid levels in comparison to their age-matched controls (Al-0/PP). On the other hand, the Al-5 group of animals failed to show any rise in the lipid parameters except HDL cholesterol. In general, the main cardioprotective function of the ‘good cholesterol’ HDL is attributed to its ability to act as both the acceptor of cholesterol from cells and as the cholesterol carrier in the reverse cholesterol transport pathway, including delivery to the liver.[36-38] This facilitates low plasma cholesterol levels in health. In our study, however, we have witnessed a significant decrease in hepatic lipid levels in both Al-5 and Al-10 exposed YA rats only and not in PP rats compared to control groups. This inverse finding in the liver and plasma lipid levels further supports that Al perturbs the lipid metabolism mainly in the YA rats, indicating that Al might lead to dyslipidaemia due to its metalloestrogenic activity. Remarkably, plasma HDL cholesterol of the Al-5/PP rats was the only parameter that showed a significant increase. Similar to the association of elevated HDL in PP obese children,[39] the increased gain in BW in the Al-5 group of PP female rats might have some correlation.
Alterations in lipid profile with hypertriglyceridemia in Al-exposed animals were reported[40,41] that ratified the current observations of raised plasma TG levels in Al-5 and Al-10 groups of YA female rats. However, this present report unfolded that the same level of Al exposure for the same duration failed to replicate similar changes in plasma TG levels of PP female rats. Similarly, PP female rats also demonstrated resistance towards changes in total plasma cholesterol level in response to Al-exposures, while YA female rats were not. Studies suggested that interaction between Al and the surface of HDL causes changes in the physicochemical properties of the lipoprotein monolayer.[42] Interaction between the negatively charged proteins with the Al3+ caused its binding to phosphate groups in the phospholipid monolayer at the HDL surface.[43] These unusual reactions might be responsible for the observed plasma and liver HDL cholesterol levels in Al-exposed animals.
Obstructions of mitochondrial functions in Al-insulted HepG2 cells indicated a disrupted Tricarboxylic acid cycle (TCA) cycle, limiting the adenosine triphosphate production, in turn, with an increase in lipid production.[44] Thus, Al might be involved in changes in lipid parameters by both direct and indirect processes, as Al-induced alterations in activities of enzymes involved in the synthesis, translocation, as well as peroxidation, also cannot be ruled out. Further to substantiate the finding of our preliminary report, the enzymatic involvement in lipid metabolism after acute Al exposure in female rats in and around puberty must be explored.
Lipid ratios and AI are used to correlate dyslipidaemia with cardiovascular risks. Increases in CRI-I, CRI-II, AC and AIP were noted in Al-exposed groups of YA female rats in comparison to respective groups of PP female rats. These indicated the role of oestrogen in Al-induced alterations in lipid profile and cardiovascular risk factors. In this context, it is worth noting that Al is a ME[4,45-47] and its involvement in oestrogen-mediated mechanisms had been suggested for long.[48,49] Mainly, these studies show how Al interferes with the oestrogen receptor signalling. Moreover, alterations in lipid profile in women with menopause[50] and its correction with oestrogen therapy[51] had already suggested the crucial role of oestrogen in lipid metabolism and dyslipidaemia. Experiments with ovariectomised rats also confirmed the role of oestrogen in liver lipid homeostasis.[52,53]
The present study sheds light on the Al-induced dyslipidaemia in YA female rats. In addition, mostly unperturbed lipid parameters, as well as lipid ratios in PP female rats, indicate the metalloestrogenic property of Al. However, the possible role of Al as ME particularly in the context of lipid homeostasis is not fully understood. Nevertheless, the study evinced the contribution of oestrogen in Al toxicity, even in acute exposure. Future studies should be directed towards unfolding the enzymatic mechanism of the metalloestrogenic property of Al, contributing to dyslipidaemia.
CONCLUSION
Variations in lipid parameters of plasma and liver observed in the current experiment suggest that Al possibly interacts differentially with the lipid metabolism depending on the reproductive age of the female rats. This significant finding is the first of its kind where Al exposure has been studied in female rats of two age groups (PP and YA), to reveal the interaction between Al and the oestrogen axis. In general, distinct oestrogen statuses in virtue of their pubertal state related to lipid metabolism have been documented.[14,54] The result reported here sheds light on the influence of oestrogen status on the Al-induced alterations in lipid profile parameters and cardiovascular risk factors. This is the first report of acute Al exposure in PP and YA female rats suggesting oestrogen’s involvement in Al-induced dyslipidaemia.
Acknowledgement
We are indebted to Prof. S Sircar, Ex-HOD, Department of Physiology, AIIMS, Jodhpur for his constant support during the entire course of the study.
Declaration of patient consent
Patient’s consent not required as there are no patients in this study.
Conflicts of interest
There are no conflicts of interest.
Financial support and sponsorship
Nil.
References
- Exposure of man to environmental aluminium--an exposure commitment assessment. Sci Total Environ. 1986;52:65-82.
- [CrossRef] [PubMed] [Google Scholar]
- Aluminium speciation in biology In: Chadwick DJ, Whelan J, eds. Novartis Foundation Symposia. Chichester, UK: John Wiley and Sons. Ltd.; 2007. p. :5-25. Available from: https://onlinelibrary.wiley.com/doi/10.1002/9780470514306.ch2 [Last accessed on 2022 May 17]
- [Google Scholar]
- Aluminium-A potentiator of multiple evils. Ind J Physiol Pharmacol. 2019;63:259-62.
- [Google Scholar]
- Aluminum-induced mitochondrial dysfunction leads to lipid accumulation in human hepatocytes: A link to obesity. Cell Physiol Biochem. 2007;20:627-38.
- [CrossRef] [PubMed] [Google Scholar]
- The putative role of environmental aluminium in the development of chronic neuropathology in adults and children. How strong is the evidence and what could be the mechanisms involved? Metab Brain Dis. 2017;32:1335-55.
- [CrossRef] [PubMed] [Google Scholar]
- The disruption of l-carnitine metabolism by aluminum toxicity and oxidative stress promotes dyslipidemia in human astrocytic and hepatic cells. Toxicol Lett. 2011;203:219-26.
- [CrossRef] [PubMed] [Google Scholar]
- Lipid profile of women qualifying for hypolipidaemic treatment. Acta Cardiol. 2006;61:440-5.
- [CrossRef] [PubMed] [Google Scholar]
- Heart disease and stroke statistics--2011 update: A report from the American heart association. Circulation. 2011;123:e18-209.
- [Google Scholar]
- Coronary heart disease mortality declines in the United States from 1979 through 2011: Evidence for stagnation in young adults, especially women. Circulation. 2015;132:997-1002.
- [CrossRef] [PubMed] [Google Scholar]
- Sex and age differences in lipoprotein subclasses measured by nuclear magnetic resonance spectroscopy: The Framingham Study. Clin Chem. 2004;50:1189-200.
- [CrossRef] [PubMed] [Google Scholar]
- Sex hormones and sex chromosomes cause sex differences in the development of cardiovascular diseases. Arterioscler Thromb Vasc Biol. 2017;37:746-56.
- [CrossRef] [PubMed] [Google Scholar]
- Changes in fasting lipids during puberty. J Pediatr. 2016;170:199-205.
- [CrossRef] [PubMed] [Google Scholar]
- Sex hormones drive changes in lipoprotein metabolism. iScience. 2021;24:103257.
- [CrossRef] [PubMed] [Google Scholar]
- Energy metabolism and fertility: A balance preserved for female health. Nat Rev Endocrinol. 2014;10:13-23.
- [CrossRef] [PubMed] [Google Scholar]
- Influence of puberty timing on adiposity and cardiometabolic traits: A Mendelian randomisation study. PLoS Med. 2018;15:e1002641.
- [CrossRef] [PubMed] [Google Scholar]
- Endocrine disrupters and human puberty. Int J Androl. 2006;29:264-71.
- [CrossRef] [PubMed] [Google Scholar]
- Prenatal lead exposure in the rat during the third week of gestation: Long-term behavioral, physiological, and anatomical effects associated with reproduction. Toxicol Appl Pharmacol. 1991;110:206-15.
- [CrossRef] [PubMed] [Google Scholar]
- Estrogen-like endocrine disrupting chemicals affecting puberty in humans--A review. Med Sci Monit. 2009;15:RA137-45.
- [Google Scholar]
- Estrogenic endocrine disrupting chemicals influencing NRF1 regulated gene networks in the development of complex human brain diseases. Int J Mol Sci. 2016;17:2086.
- [CrossRef] [PubMed] [Google Scholar]
- Low doses of bisphenol a induce gene expression related to lipid synthesis and trigger triglyceride accumulation in adult mouse liver. Hepatology. 2012;55:395-407.
- [CrossRef] [PubMed] [Google Scholar]
- Cadmium's disguise dupes the estrogen receptor. Nat Med. 2003;9:1000-1.
- [CrossRef] [PubMed] [Google Scholar]
- Estrogen-Like activity of metals in Mcf-7 breast cancer cells. Endocrinology. 2003;144:2425-36.
- [CrossRef] [PubMed] [Google Scholar]
- In utero and peripubertal metals exposure in relation to reproductive hormones and sexual maturation and progression among girls in Mexico City. Environ Res. 2019;177:108630.
- [CrossRef] [PubMed] [Google Scholar]
- Dissimilar anxiety-like behavior in prepubertal and young adult female rats on acute exposure to aluminium. Cent Nerv Syst Agents Med Chem. 2021;21:187-94.
- [CrossRef] [PubMed] [Google Scholar]
- The effects of aqueous ginger extract on aluminium chloride (AlCl3) induced alteration in lipid profile of male wister rats. Int J of Basic Clin Pharmacol. 2017;6:2587-90.
- [CrossRef] [Google Scholar]
- The effects of low dose aluminum on hemorheological and hematological parameters in rats. Arch Toxicol. 2007;81:11-7.
- [CrossRef] [PubMed] [Google Scholar]
- Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin Chem. 1972;18:499-502.
- [CrossRef] [PubMed] [Google Scholar]
- A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 1957;226:497-509.
- [CrossRef] [PubMed] [Google Scholar]
- Evaluation of dyslipidemia, lipid ratios, and atherogenic index as cardiovascular risk factors among semi-urban dwellers in Nigeria. Ann Afr Med. 2016;15:194-9.
- [CrossRef] [PubMed] [Google Scholar]
- Past : Paleontological statistics software package for education and data analysis. Palaenotol Electron. 2001;4:1-9.
- [Google Scholar]
- Lifelong feeding of a high aluminum diet to mice. Toxicology. 2000;150:107-17.
- [CrossRef] [PubMed] [Google Scholar]
- Effects of aluminium chloride exposure on the histology of lungs of wistar rats. J Appl Pharm Sci2013;. ;3:108-12.
- [CrossRef] [Google Scholar]
- HDL and reverse cholesterol transport. Circ Res. 2019;124:1505-18.
- [CrossRef] [PubMed] [Google Scholar]
- High-density lipoproteins: Nature's multifunctional nanoparticles. ACS Nano. 2016;10:3015-41.
- [CrossRef] [PubMed] [Google Scholar]
- Role of plasma lecithin: Cholesterol acyltransferase in the metabolism of high density lipoproteins. J Lipid Res. 1966;7:638-48.
- [CrossRef] [PubMed] [Google Scholar]
- Triglycerides-to-HDL ratio as a new marker of endothelial dysfunction in obese prepubertal children. Eur J Endocrinol. 2013;170:173-80.
- [CrossRef] [PubMed] [Google Scholar]
- Study of lipid profile and parieto-temporal lipid peroxidation in AlCl3 mediated neurotoxicity, modulatory effect of fenugreek seeds. Lipids Health Dis. 2012;11:16.
- [CrossRef] [PubMed] [Google Scholar]
- Protective effects of Aloe vera extract on aluminium sulphate induced alterations in serum lipid profile of male albino rats, Rattus norvegicus. Biosci Biotechnol Res Commun. 2018;11:727-33.
- [CrossRef] [Google Scholar]
- Effect of aluminium on lipid peroxidation of human high density lipoproteins. Free Radic Res. 2003;37:515-21.
- [CrossRef] [PubMed] [Google Scholar]
- TLQP-21, a VGF-derived peptide, increases energy expenditure and prevents the early phase of diet-induced obesity. Proc Natl Acad Sci. 2006;103:14584-9.
- [CrossRef] [PubMed] [Google Scholar]
- Mechanism of aluminum-induced inhibition of hepatic glycolysis: Inactivation of phosphofructokinase. J Pharmacol Exp Ther. 1990;254:301-5.
- [Google Scholar]
- Potential interference of aluminum chlorohydrate with estrogen receptor signaling in breast cancer cells. J Mol Biochem. 2018;7:1-13.
- [Google Scholar]
- Neurotoxic effects of aluminum are associated with its interference with estrogen receptors signaling. Neurotoxicology. 2020;77:114-26.
- [CrossRef] [PubMed] [Google Scholar]
- The potential for estrogen disrupting chemicals to contribute to migration, invasion and metastasis of human breast cancer cells. J Cancer Metastasis Treat. 2019;5:58.
- [CrossRef] [Google Scholar]
- Metalloestrogens: An emerging class of inorganic xenoestrogens with potential to add to the oestrogenic burden of the human breast. J Appl Toxicol. 2006;26:191-7.
- [CrossRef] [PubMed] [Google Scholar]
- Environmental oestrogens and breast cancer: Evidence for combined involvement of dietary, household and cosmetic xenoestrogens. Anticancer Res. 2010;30:815-27.
- [Google Scholar]
- Clinical review 91: Female sex hormones and cardiovascular disease in women. J Clin Endocrinol Metab. 1997;82:3913-8.
- [CrossRef] [PubMed] [Google Scholar]
- Estrogen reduces lipid content in the liver exclusively from membrane receptor signaling. Sci Signal. 2013;6:ra36.
- [CrossRef] [PubMed] [Google Scholar]
- Role of estrogens in the regulation of liver lipid metabolism In: Mauvais-Jarvis F, ed. Sex and Gender Factors Affecting Metabolic Homeostasis, Diabetes and Obesity. Cham: Springer International Publishing; 2017. p. :227-56. Available from: https://link.springer.com/10.1007/978-3-319-70178-3_12 [Last accessed on 2022 May 19]
- [CrossRef] [PubMed] [Google Scholar]
- Age, maturation and serum lipid parameters: Findings from the German health survey for children and adolescents. BMC Public Health. 2019;19:1627.
- [CrossRef] [PubMed] [Google Scholar]




