
Translate this page into:
Silica nanoparticles: Chemical synthesis and acute neurotoxic study in Wistar rat
*Corresponding author: Placheril J. John, Department of Zoology, University of Rajasthan, Jaipur, Rajasthan, India. placheriljohn@gmail.com
-
Received: ,
Accepted: ,
How to cite this article: Yadav S, Nirmal NK, Gupta H, John PJ. Silica nanoparticles: Chemical synthesis and acute neurotoxic study in Wistar rat. Indian J Physiol Pharmacol. 2024;68:33-41. doi: 10.25259/IJPP_139_2023
Abstract
Objectives:
In this study, silica nanoparticles (SNPs) were synthesized through the sol-gel approach, characterized using X-ray diffraction and examined for their potential to induce acute neurotoxicity in rats.
Materials and Methods:
The average particle size was confirmed to be 110 ± 5 nm through scanning electron microscope (SEM) analysis. An intraperitoneal injection of 25, 50, and 100 mg/kg of SNPs was administered twice over 24 h to rats, followed by autopsies 24 h later. Brain regions (cerebellum, frontal cortex, hippocampus, and corpus striatum) were analyzed for changes in antioxidant enzyme activity (superoxide dismutase [SOD], catalase [CAT], glutathione-s-transferase [GST], glutathione peroxidase [GPx], glutathione reductase [GR] and thiobarbituric acid reactive substance [TBARS] levels) and neurotransmitter concentrations (acetylcholinestrase [AChE], dopamine, serotonin and nor-epinephrine).
Results:
The activity of SOD, CAT and GST increased significantly in the high dose group, specifically in the hippocampus and corpus striatum. No significant change in GR activity was observed in any dose group. Significant alterations were observed in neurotransmitter concentrations in hippocampus and corpus striatum in rats treated with high dose.
Conclusion:
Overall, the outcomes derived from our research suggest that SNPs lead to dose-dependent oxidative harm and neurotoxic effects in different regions of brain.
Keywords
Silica nanoparticles
Wistar rat
Antioxidant
Neurotransmitter

INTRODUCTION
Silica nanoparticles (SNPs) are increasingly being used in various consumer, environmental and bio-medical applications.[1,2] Importantly, SNPs have already entered human clinical trials as diagnosis, drug delivery vehicles and agents for photothermal ablation therapy.[3] The risk of exposure to biological systems, including humans, is thus raised. Human exposure to these particles may take place through inhalation, ingestion, dermal and injection routes including exposure at workplaces such as research laboratories, automobile, aerospace factories, gun industries or pharmaceutical industries.[4]
A limited number of studies have measured exposure and potential risks for humans. McCarthy et al. (2012) reported inflammation, oxidative damage, and death of human lung submucosal cells when exposed to amorphous SNPs in vitro for 24 h.[5] Similarly, SNP exposure leads to significant oxidative stress followed by apoptosis in cultured cardiomyocytes.[6] An in vitro study conducted using Wistar rats examined the toxicity of SNPs after inhalation exposure. There was clear evidence of chronic kidney inflammation and fibrosis even after stopping the exposure. Focal lesions in the lungs were also reported.[7]
Few studies have examined the effects of SNP exposure on the brain. Wu et al. (2011) demonstrated that intranasally instilled SNPs entered the brain.[8] The particles were associated with increased inflammation and oxidative damage in the striatum area of the brain in treated rats.[8] In another study, male mice were exposed to silica nanomaterials through intranasal instillation once per day for 81 days. SNPs cause elevated tau phosphorylation, neuroinflammation, and neurodegeneration-like features in the medial prefrontal cortex and the hippocampus.[9] Clearly, there is evidence that SNPs can potentially harm nervous tissue through several mechanisms. However, a complete understanding of neurotoxic effects and the underlying mechanism is still lacking.[10]
The current investigation, therefore, aims to fill this knowledge gap. Amorphous, mesoporous, and spherical SNPs were synthesised using a sol-gel chemical procedure. Subsequently, adult Wistar rats were acutely exposed to varying concentrations of SNPs through intraperitoneal administration. General health, oxidative stress parameters, neurotransmitter concentrations, and histopathological changes were examined in four specific brain regions, namely the cerebellum, frontal cortex, hippocampus, and corpus striatum of control and treated animals. The present study is expected to provide insights that will help to understand the potential risks associated with SNP exposure in various biomedical, occupational, and environmental settings.
MATERIALS AND METHODS
Materials
Tetraethyl orthosilicate (TEOS) (TEOS ≥99.0% GC M.W: 208.33 g/moL, d: 0.933 g/mL) and mesitylene (M.W: 120.19 g/moL, d = 0.864 g/mL, 98%) were procured from Sigma-Aldrich, and N-cetyl-N,N,N-try methylammonium bromide (CTAB, M.W: 364.45, 99%) and sodium hydroxide (NaOH) were procured from HiMedia Laboratories (Mumbai, India) for SNPs preparation. Ultrapure water (Direct-Q® Water Purification System, Merck Millipore) was used all over synthesis to avoid metal impurities. Analytical-grade chemicals were used in all the experiments.
Synthesis of SNPs
A sol-gel chemical method was used to synthesise spherical and mesoporous SNPs in the laboratory according to the procedure described by Rahman and Padavettan (2012).[11] Briefly, the methodology involves the synthesis of SNPs using CTAB as a surfactant and mesitylene as a pore-size modifier [Figure 1]. The process involves dissolving CTAB in water, adjusting the pH to between 11 and 12 using NaOH, adding mesitylene, and then adding TEOS dropwise while stirring at 80°C. The resulting white precipitate is hot-filtered, washed with water and acidic ethanol to remove the CTAB template, and dried under a light lamp. The yield of the dried SNP powder was 1.006 g.

- Synthesis of silica nanoparticles (SNPs) in the study. A sol-gel chemical method was used to synthesise spherical and mesoporous SNPs in the laboratory according to the procedure described by Rahman and Padavettan. CTAB: Detyltrimethylammonium bromide, NaOH: Sodium hydroxide, TEOS: Tetraethyl orthosilicate, rpm: Revolutions per minute.
Particle characterisation
The amorphous structure of spherical SNPs was analysed using X-ray diffraction with a PANalytical X-ray Diffractometer, using Cu Kα radiation with a wavelength of 1.54060 Å. The analysis was conducted in the 2θ range of 10–90° at a scan speed of 0.4°/min to confirm the structural properties of SNPs. The optical properties of the SNPs were analysed using an ultraviolet–visible spectroscopy (UV-Vis) spectrophotometer operating in the 200–700 nm wavelength range, with a resolution of 2 nm. The particle size and distribution of SNPs were measured using dynamic light scattering with a Malvern Zetasizer instrument, with 80 accumulation times per measurement. The zeta potential of the SNPs was measured using electrophoretic light scattering with the same instrument to determine the reason for agglomeration. The size distribution and morphology of SNPs were analysed using SEM images obtained with a 30 kV EVO-18 Carl Zeiss Scanning Electron Microscope. The diameter of 100 randomly selected SNPs was measured to determine the size distribution.
Animal handling
In this study, healthy male Wistar rats with a body weight of 120 ± 10 g an age of 6–8 weeks, and proven fertility, were used. They were housed under standard laboratory conditions, with a temperature of 25 ± 2°C and relative humidity of 35 ± 5%, with a 12-h light and 12-h dark photoperiod, in polypropylene cages with clean wooden shavings as bedding. Animals were fed a standard diet. The animal handling and care followed the guidelines set by the Committee for the Purpose of Control and Supervision on Experiments on Animals in New Delhi, India. The experiment had the consent of the departmental animal ethics committee (UDZ/IAEC/1/020-CPCSEA, dated 13 July 2017).
Experimental schedule
Rats were administered SNPs at doses of 25, 50, and 100 mg/kg twice over 24 h through intraperitoneal injection as per the guidelines set by the Committee for the Purpose of Control and Supervision on Experiments on Animals in New Delhi, India. The rats were divided into four groups (6 animals/group) including a control group (ultra-pure water as the vehicle), a low dose group (25 mg/kg), a moderate dose group (50 mg/kg), and a high dose group (100 mg/kg). After 24 h, the rats were autopsied and brain sub-regions (frontal cortex, cerebellum, hippocampus, and corpus striatum) were analysed for changes in antioxidant enzyme activity and biochemical parameters to assess acute neurotoxicity.
Analysis of antioxidant enzymes activity and thiobarbituric acid reactive substance (TBARS) level assessment
The rats were euthanised and autopsied to isolate their brains, which were weighed and divided into four regions: Cerebellum, frontal cortex, hippocampus, and corpus striatum. The brain regions were homogenized in a buffer solution and centrifuged to obtain a 10% homogenate. This homogenate was used to measure the activity of various antioxidant enzymes and neurotransmitters. The activity of acetylcholinesterase (AChE) was also assessed.
Enzymatic activities of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), glutathione-s-transferase (GST) and glutathione reductase (GR) were assessed as per the protocols established by Marklund and Marklund, Aebi, Paglia and Valentine, Habig et al. and Bergmeyer and Garvehn, respectively.[12-16] TBARS concentration was measured in brain tissue homogenates using the protocol of Ohkawa et al.[17] Concentrations of dopamine, serotonin and norepinephrine were measured by the methods prescribed by Jacobowitz and Richardson.[18] The activity of AChE enzyme was evaluated following the method given by Ellman et al.[19]
Statistical analysis
All the data were represented as mean ± SD and statistically analysed by GraphPad Prism (v. 8.0.2). One-way analysis of variance followed by Dunnett’s multiple comparisons test was utilised to test the statistical difference between mean values from control and treated groups. P < 0.05 was considered as statistically significant.
RESULTS
Particle characterisation
X-ray diffraction and UV-Vis spectrum
UV-Vis spectrophotometer (Varian) was used to analysis the optical properties of colloidal solution of SNPs in the region of 200–700 nm operating at a resolution of 2 nm.[20] Absorption peak λ max was obtained at 270 nm. Figure 2a shows the absorption spectra of synthesised SNPs. The results, shown in Figure 2b, reveal the non-crystalline and amorphous nature of the SNPs through the presence of a broad peak at 2θ = 22° and a wide hump between 16° and 30°, indicating the absence of any crystalline structures. These findings are consistent with previous studies on SNPs reported in the literature.[21,22]

- (a) Ultraviolet-visible spectrum of silica nanoparticles. (b) Shows X-ray diffraction pattern of silica nanoparticles (SNPs) that confirms the amorphous structure of SNPs.
Zeta potential and size distribution by intensity
The measurement of zeta potential of SNPs was recorded to obtain the reason for agglomeration.[23] Malvern Zetasizer instrument with electrophoretic light scattering was used to measure the zeta potential. The measurements were repeated 5 times and average values were recorded. The graph shows good stability of synthesised particles and very less tendency to form aggregates [Figure 3a]. The intensity-weighted average particle size (nm) and particle distribution of SNPs were measured. The hydrodynamic size obtained was between 300 and 600 nm [Figure 3b].

- (a) Zeta potential of silica nanoparticle. (b) The size distribution curve of silica cluster nanoparticles. b
Scanning electron microscopy
SEM was used to find the average particle size and the morphology of the SNPs using by 30 kV EVO-18 Carl Zeiss SEM. The SEM images as shown in [Figures 4a and b]. SNPs were observed in the size range of 80–115 nm. The SEM micrograph indicates that the synthesised SNPs are largely spherical in shape and size. However, some particles had non-spherical shapes like elongated, ellipsoidal, larger aggregates. The SEM analysis has revealed that the average particle size is 113 nm.

- (a and b) SEM images of different-sized silica nanoparticles (SNPs). The particle size of SNPs was recorded in between 80 and 115 nm range. The size distribution curve shows that the synthesised SNPs had a narrow size range.
Oxidative stress markers in the brain
The study evaluated the changes in the activities of various antioxidant enzymes, such as SOD, CAT, GST, GPx, and GR as well as the level of TBARS in four sub-regions of the brain: Cerebellum, frontal cortex, hippocampus, and corpus striatum. The results showed an increase in the activities of CAT, SOD and GST in hippocampus and corpus striatum sub-regions of the brain of animals exposed to high doses of SNPs compared to the control group after 24 h of exposure [Figures 5a-c]. GR activity showed statistically insignificant changes in all treated groups compared to the control group, in all the brain sub-regions exposed to SNPs in any dose group, as shown in Figure 5d. GPx and TBARS levels also increased in the hippocampus and corpus striatum sub-regions [Figures 5e, f and Table 1]. Antioxidant enzyme activities remained statistically comparable to control in the cerebellum and frontal cortex regions.

- The specific activity of antioxidant enzymes in different brain sub-regions (cerebellum, frontal cortex, hippocampus and corpus striatum) of rats treated with control and different doses of silica nanoparticles. (a) Catalase (CAT). (b) Superoxide dismutase (SOD). (c) GlutathioneS-transferase (GST). (d) Glutathione reductase (GR). (e) Glutathione peroxidase (GPx). (f) Concentration of thiobarbituric acid reactive substances (TBARS). Each group consisted of 6 animals and the values are expressed as mean±SD. The data were analysed using one-way analysis of variance followed by Dunnett’s multiple comparison test (*P ≤ 0.05). SOD (1 unit is defined as the amount of enzyme inhibiting the rate of autoxidation of pyrogallol by 50%), CAT (1 unit is equal to 1 mmol H2O2 decomposed per min per mg protein), GST (μ mole G-SDNB formed per min per mg protein), GR (1 unit equals to nmol NADPH oxidised per min per mg protein), GPx (1 unit is defined as nmol NADPH oxidised/min/mg protein), TBARS (1 unit is equal to 1 nmol TBARS formed/h/mg protein). C: Control group, LD: Low dose group, MD: Moderate dose group, HD: High dose group.
| Parameter | Group | Cerebellum | Frontal cortex | Hippocampus | Corpus striatum |
|---|---|---|---|---|---|
| CAT | Control | 0.99±0.09 | 0.9±0.09 | 0.92±0.11 | 0.85±0.1 |
| L.D. | 1.13±0.15 | 0.92±0.12 | 1.09±0.14 | 0.93±0.1 | |
| M.D. | 1.16±0.14 | 0.96±0.14 | 1.08±0.16 | 0.98±0.14 | |
| H.D. | 1.18±0.15 | 1.05±0.17 | 1.16±0.13* | 1.16±0.17* | |
| SOD | Control | 126±4.85 | 103±5.54 | 111±5.2 | 104±4.3 |
| L.D. | 132±6.32 | 106±6.61 | 115±6.96 | 108±7.63 | |
| M.D. | 136±10.25 | 108±10.2 | 158±10.22* | 120±12.23 | |
| H.D. | 140±16.32 | 110±16.34 | 165±16.36* | 142±16.4* | |
| GST | Control | 20.56±1.23 | 21.26±1.99 | 22.89±1.34 | 22.65±1.48 |
| L.D. | 21.56±1.56 | 22.9±1.68 | 24.67±1.54 | 24.28±1.62 | |
| M.D. | 22.08±1.36 | 23.01±1.12 | 25.01±1.3 | 25.1±1.85 | |
| H.D. | 23.04±1.71 | 23.75±1.98 | 27.46±1.7* | 27.26±1.8* | |
| GR | Control | 3.26±0.22 | 2.9±0.32 | 3.05±0.58 | 2.92±0.7 |
| L.D. | 3.45±0.32 | 2.95±0.84 | 3.54±0.57 | 3.58±0.45 | |
| M.D. | 3.58±0.5 | 3.09±0.43 | 3.89±0.65 | 3.75±0.71 | |
| H.D. | 3.58±0.64 | 3.11±0.37 | 3.91±0.56 | 3.8±0.55 | |
| GPx | Control | 21.84±2.44 | 25.56±2.42 | 25.56±2.4 | 25.12±2.45 |
| L.D. | 22.86±2.28 | 26.65±2.34 | 28.36±2.3 | 27.36±2.2 | |
| M.D. | 24.89±2.56 | 28.12±2.48 | 29.02±2.5 | 28.13±2.53 | |
| H.D. | 24.8±2.42 | 29.05±2.45 | 31.64±2.51* | 31.86±2.15* | |
| TBARS | Control | 3.85±0.36 | 3.34±0.54 | 2.09±0.68 | 3.06±0.95 |
| L.D. | 3.96±0.65 | 3.55±0.68 | 2.56±0.89 | 3.56±0.79 | |
| M.D. | 4.02±0.54 | 3.86±0.95 | 2.87±0.65 | 4.56±0.92 | |
| H.D. | 4.11±0.67 | 4.02±0.64 | 3.17±0.7* | 5.78±0.95* |
CAT: Catalase, SOD: Superoxide dismutase, GST: Glutathione S-transferase, GR: Glutathione reductase, GPx: Glutathione peroxidase, TBARS: Thiobarbituric acid reactive substances, L.D.: Low dose (25 mg/kg), M.D.: Medium dose (50 mg/kg), H.D.: High dose (100 mg/kg), SNPs: Silica nanoparticles. Values marked with an asterisk (*) represent a significant difference in the antioxidant enzyme activity as compared to control. The presented values are mean±standard deviation
Neurotransmitters level in brain
The levels of serotonin and nor-epinephrine were observed to have a significant decline in high dose group in hippocampus and corpus striatum compared to the control group [Figures 6a, b and Table 2]. SNPs did not alter the dopamine levels of various brain sub-regions in any dose group as shown in figure 6c. Activity of acetylcholine esterase enzyme showed significant decline in high dose group in hippocampus and corpus striatum of treated rats [Figure 6d]. Neurotransmitter levels remained statistically comparable to control in cerebellum and frontal cortex regions.
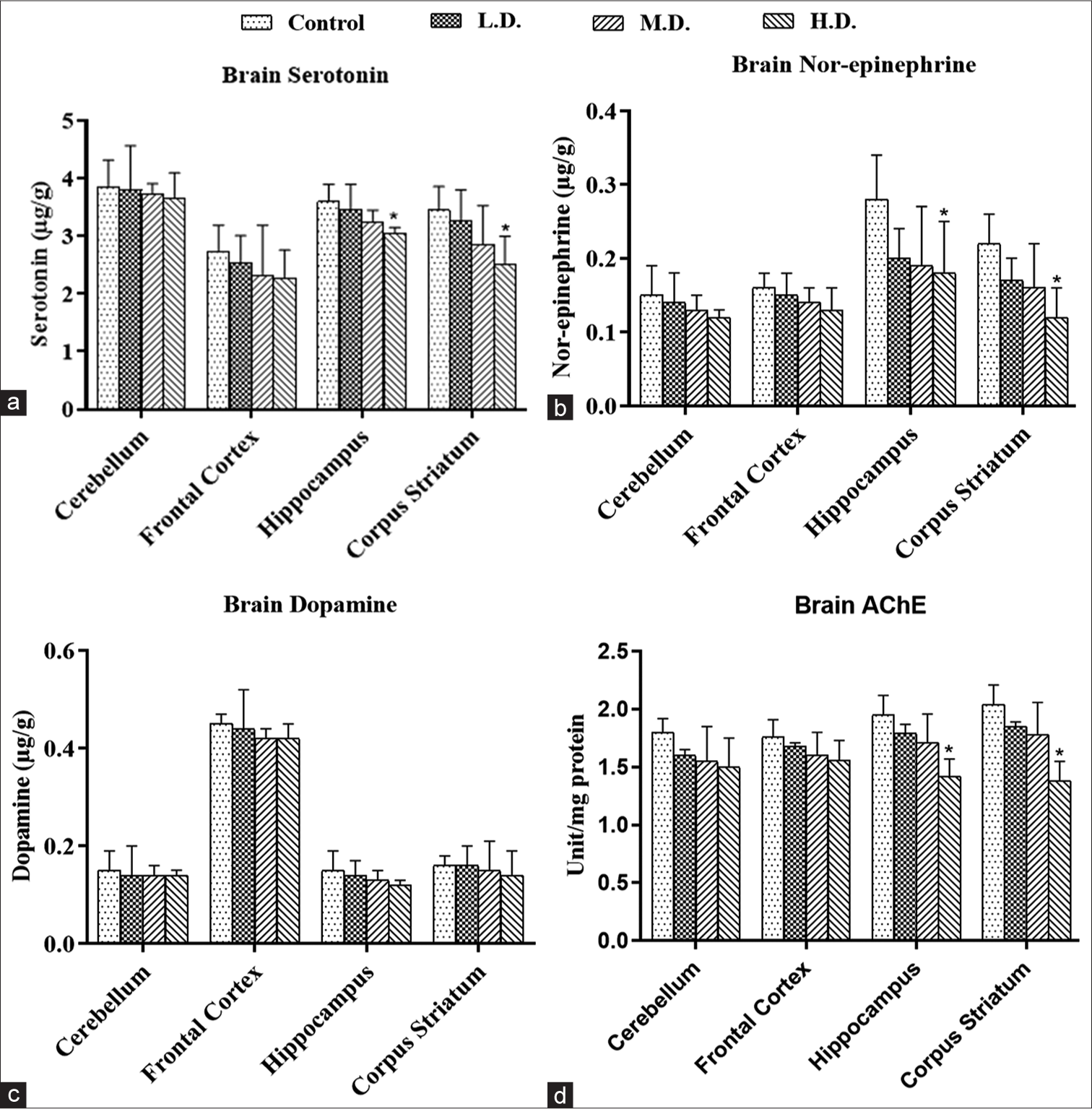
- (a-d) The concentration of neurotransmitters in different brain sub-regions (cerebellum, frontal cortex, hippocampus and corpus striatum) of control and silica nanoparticles-treated rats. (a) Serotonin. (b) Norepinephrine. (c) Dopamine (in μg/g). (d) The specific activity of acetylcholinesterase (in μ mol substrate hydrolysed/min/mg protein). Each group had 6 animals, and the values are presented as mean±SD. The data were analysed using one-way analysis of variance followed by Dunnett’s multiple comparison test (*P≤0.05). C: Control group, LD: Low dose group, MD: Moderate dose group, HD: High dose group.
| Parameter | Group | Cerebellum | Frontal cortex | Hippocampus | Corpus striatum |
|---|---|---|---|---|---|
| Serotonin | Control | 3.85±0.47 | 2.74±0.45 | 3.6±0.3 | 3.45±0.41 |
| L.D. | 3.8±0.77 | 2.54±0.47 | 3.45±0.45 | 3.26±0.54 | |
| M.D. | 3.74±0.17 | 2.32±0.87 | 3.25±0.2 | 2.86±0.67 | |
| H.D. | 3.65±0.45 | 2.28±0.48 | 3.05±0.1* | 2.5±0.5* | |
| Nor-epinephrine | Control | 0.15±0.04 | 0.16±0.02 | 0.28±0.06 | 0.22±0.04 |
| L.D. | 0.14±0.04 | 0.15±0.03 | 0.2±0.04 | 0.17±0.03 | |
| M.D. | 0.13±0.02 | 0.14±0.02 | 0.19±0.08 | 0.16±0.06 | |
| H.D. | 0.12±0.01 | 0.13±0.03 | 0.18±0.07* | 0.12±0.04* | |
| Dopamine | Control | 0.15±0.04 | 0.45±0.02 | 0.15±0.04 | 0.16±0.02 |
| L.D. | 0.14±0.06 | 0.44±0.08 | 0.14±0.03 | 0.16±0.04 | |
| M.D. | 0.14±0.02 | 0.42±0.02 | 0.13±0.02 | 0.15±0.06 | |
| H.D. | 0.14±0.01 | 0.42±0.03 | 0.12±0.01 | 0.14±0.05 | |
| AChE | Control | 1.8±0.12 | 1.76±0.15 | 1.95±0.17 | 2.04±0.17 |
| L.D. | 1.6±0.05 | 1.68±0.03 | 1.79±0.08 | 1.85±0.04 | |
| M.D. | 1.55±0.3 | 1.6±0.2 | 1.71±0.25 | 1.78±0.28 | |
| H.D. | 1.5±0.25 | 1.56±0.17 | 1.42±0.15* | 1.38±0.17* |
AChE: Acetylcholinesterase, L.D.: Low dose (25 mg/kg), M.D.: Medium dose (50 mg/kg), H.D.: High dose (100 mg/kg), SNPs: Silica nanoparticles. Values marked with an asterisk (*) represent a significant difference in the neurotransmitter levels or AChE activity in the high dose group when compared to the control
DISCUSSION
In this study, we have demonstrated that SNPs with an average size of 113 nm can breach the blood–brain barrier, resulting in oxidative stress and brain inflammation in Wistar rats. For instance, Wu et al. (2011) demonstrated that intranasally instilled SNPs accumulate in the corpus striatum of treated animals. SNPs induced oxidative stress and downregulation of tyrosine hydroxylase protein. Adverse effects were observed on dopaminergic neurons in the study.[8] Similarly, Liu et al. (2017) have also shown the involvement of oxidative stress and Rho-kinase/JNK signalling pathways in the toxicity caused by SNPs.[24] These particles can also impact the astrocytes, which are neuroprotective and supportive of neuron functioning.[25] In our study, acute exposure (i.p.) to SNPs at 100 mg/kg body weight in rats altered the activities of antioxidant enzymes in the hippocampus and corpus striatum, consistent with the reports discussed above. Our research indicates that SNPs may permeate specific regions of the brain, particularly the hippocampus and striatum. This infiltration is followed by the generation of oxidative stress, as suggested by our findings.
Oxidative stress is a major contributing factor in SNPs mediated cellular damage, especially, within the brain for two reasons. First, neurons are dependent mainly upon oxidative phosphorylation for energy, and second, they are exposed to more amount of oxygen.[26] Zhou et al. (2016) exposed PC12 cells, an in vitro model for dopaminergic neurons, with mesoporous SNPs. Analysis showed that the particles induced the production of ROS and malondialdehyde.[27] The exposure was also associated with reduction of reduced glutathione and leakage of lactate dehydrogenase in a dose-dependent manner.[27] Yuan et al. (2022) evaluated the effects of SNPs in a transgenic mice model for Parkinson’s disease.[28] Male mice were exposed through intranasal instillation at a dose of 5 μg/μL SNPs every 2 days for a period of 90 days.
The authors clearly demonstrated the generation of oxidative stress and damage to mitochondria. These changes lead to apoptosis in neurons.[28] The results of our study are in line with these earlier findings, further reinforcing the notion that SNPs contribute significantly to oxidative stress. AChE enzyme plays a key role in regulating the levels of acetylcholine. A decrease in AChE activity is associated with neuronal damage. Boukholda et al. (2022) administered SNPs intraperitoneally to Wistar rats at doses of 25 and 100 mg/kg bw/day for a period of 28 days.[29] Their findings demonstrated elevated reactive oxygen species and a significant reduction in the activity of the AChE enzyme in the corpus striatum. The group also demonstrated extensive histological damage in the striatum region of SNP-exposed rats. The present study showed a similar decline in AChE enzyme activity in different brain sub-regions of SNPs-exposed rats.[29] Our results also demonstrated a significant decline in norepinephrine and serotonin levels in the hippocampus and striatum. We hypothesise that these alterations may be due to the disruption of redox processes within the brain tissue leading to structural damage to neuronal cells following SNPs exposure [Figure 7]. This speculation aligns with the findings of our current investigation and is further substantiated by previous studies,[30-32] which highlight the potential for SNPs to disturb redox homeostasis.
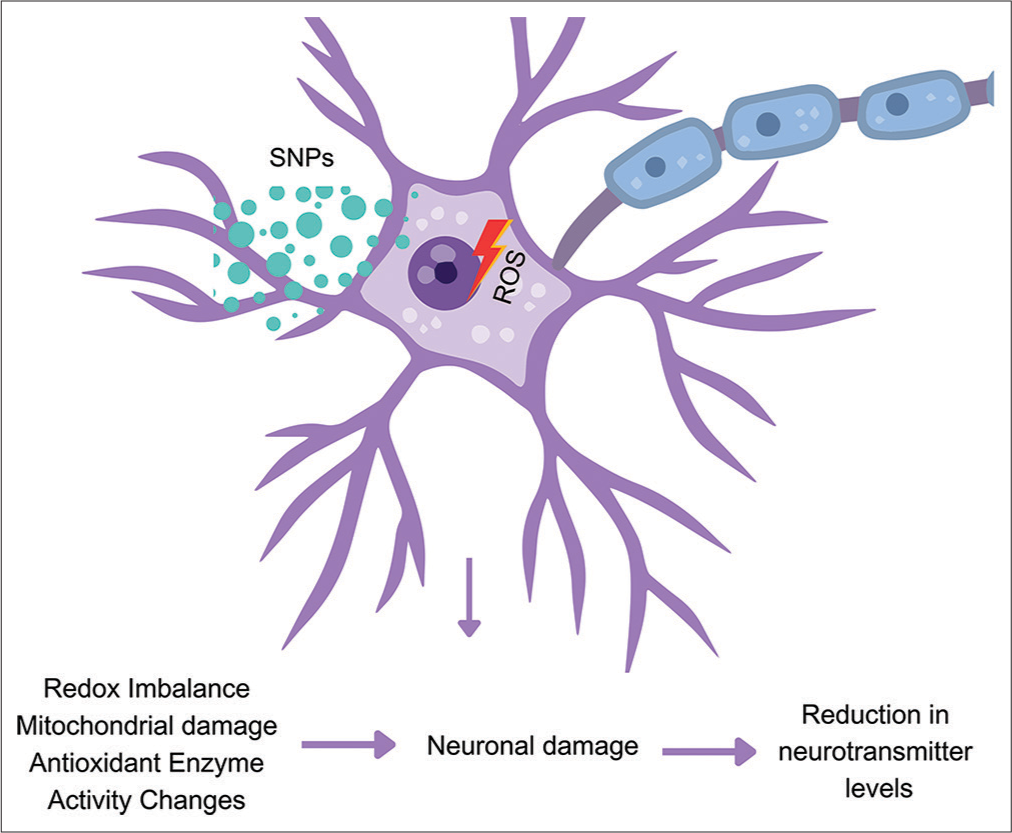
- Proposed mechanism of silica nanoparticles (SNPs) induced pathology in the brain of rats treated with intraperitoneal injections of SNPs was administered twice over 24 h. ROS: Reactive oxygen species.
CONCLUSION
The present study was designed to examine the neurotoxic effects caused by the acute exposure of SNPs to Wistar rats. SNPs utilised in the study, were synthesised utilising the sol-gel method and were amorphous and spherical in shape (110 ± 5 nm). The findings demonstrated that SNPs altered the activities of antioxidants enzymes, such as SOD, CAT, GST, GR, and GPx in the hippocampus and corpus striatum of treated rats, specifically at 100 mg/kg dose. Serotonin and norepinephrine levels also declined in these brain regions. The activity of AChE enzyme was also decreased due to the exposure. Overall, the results clearly indicate that acute exposure to SNPs induces oxidative stress and neurotoxicity in the brain. It is imperative that further studies should be undertaken to delve deeper into the in vivo toxicity of SNPs to better understand the potential risks they pose, paving the way for safer and more informed applications of these nanoparticles in the future.
Acknowledgement
The design of the procedure was planned by SY and PJJ. SNPs synthesis and their in vivo toxicity studies were accomplished by SY, NKN, and Hemant. The authors sincerely thank Dr. Kamlendra Awasthi for providing technical support in material characterisation.
Ethical approval
The author(s) declare that they have taken the ethical approval from animal ethics committee.
Declaration of patient consent
The authors certify that they have obtained all appropriate consent.
Conflicts of interest
There are no conflicts of interest.
Use of artificial intelligence (AI)-assisted technology for manuscript preparation
The authors confirm that there was no use of artificial intelligence (AI)-assisted technology for assisting in the writing or editing of the manuscript and no images were manipulated using AI.
Financial support and sponsorship
Nil.
References
- Applications of mesoporous materials as excipients for innovative drug delivery and formulation. Curr Pharm Des. 2013;19:6270-89.
- [CrossRef] [PubMed] [Google Scholar]
- Recent progress in the applications of silica-based nanoparticles. RSC Adv. 2022;12:13706-26.
- [CrossRef] [PubMed] [Google Scholar]
- Clinical translation of silica nanoparticles. Nat Rev Mater. 2021;6:1072-4.
- [CrossRef] [PubMed] [Google Scholar]
- Occupational exposure to silica nanoparticles: Evaluation of emission fingerprints by laboratory simulations. Sustainability. 2022;14:10251.
- [CrossRef] [Google Scholar]
- Mechanisms of toxicity of amorphous silica nanoparticles on human lung submucosal cells in vitro: Protective effects of fisetin. Chem Res Toxicol. 2012;25:2227-35.
- [CrossRef] [PubMed] [Google Scholar]
- Oxidative stress-mediated mitochondrial pathway-dependent apoptosis is induced by silica nanoparticles in H9c2 cardiomyocytes. Toxicol Mech Methods. 2020;30:646-55.
- [CrossRef] [PubMed] [Google Scholar]
- Inhaled silica nanoparticles cause chronic kidney disease in rats. Am J Physiol Renal Physiol. 2022;323:F48-58.
- [CrossRef] [PubMed] [Google Scholar]
- Neurotoxicity of silica nanoparticles: Brain localization and dopaminergic neurons damage pathways. ACS Nano. 2011;5:4476-89.
- [CrossRef] [PubMed] [Google Scholar]
- Silica nanoparticles induce neurodegeneration-like changes in behavior, neuropathology, and affect synapse through MAPK activation. Part Fibre Toxicol. 2018;15:28.
- [CrossRef] [PubMed] [Google Scholar]
- A toxicological profile of silica nanoparticles. Toxicol Res (Camb). 2022;11:565-82.
- [CrossRef] [PubMed] [Google Scholar]
- Synthesis of silica nanoparticles by sol-gel: Size-dependent properties, surface modification, and applications in silica-polymer nanocomposites-a review. J Nanomater. 2012;2012:132424.
- [CrossRef] [Google Scholar]
- Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem. 1974;47:469-74.
- [CrossRef] [PubMed] [Google Scholar]
- Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med. 1967;70:158-69.
- [Google Scholar]
- Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J Biol Chem. 1974;249:7130-9.
- [CrossRef] [PubMed] [Google Scholar]
- Glutathione reductase In: Methods of enzymatic analysis. New York: Academic Press; 1978. p. :574-9.
- [Google Scholar]
- Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95:351-8.
- [CrossRef] [PubMed] [Google Scholar]
- Method for the rapid determination of norepinephrine, dopamine, and serotonin in the same brain region. Pharmacol Biochem Behav. 1978;8:515-9.
- [CrossRef] [PubMed] [Google Scholar]
- A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol. 1961;7:88-95.
- [CrossRef] [PubMed] [Google Scholar]
- Study on physical properties of dispersed silica. IOP Conf Ser Mater Sci Eng. 2019;684:12011.
- [CrossRef] [Google Scholar]
- Precipitation of amorphous SiO2 particles and their properties. Braz J Chem Eng. 2011;28:89-94.
- [CrossRef] [Google Scholar]
- Effect of silica nanoparticles and BTCA on physical properties of cotton fabrics. Mater Res. 2011;14:552-9.
- [CrossRef] [Google Scholar]
- The relationship between pH and zeta potential of ~30 nm metal oxide nanoparticle suspensions relevant to in vitro toxicological evaluations. Nanotoxicology. 2009;3:276-83.
- [CrossRef] [Google Scholar]
- Blood-brain barrier dysfunction induced by silica NPs in vitro and in vivo: Involvement of oxidative stress and Rho-kinase/JNK signaling pathways. Biomaterials. 2017;121:64-82.
- [CrossRef] [PubMed] [Google Scholar]
- Astrocytes are more vulnerable than neurons to silicon dioxide nanoparticles toxicity in vitro. Toxics. 2020;8:51.
- [CrossRef] [PubMed] [Google Scholar]
- Implications for blood-brain-barrier permeability, in vitro oxidative stress and neurotoxicity potential induced by mesoporous silica nanoparticles: Effects of surface modification. RSC Adv. 2016;6:2800-9.
- [CrossRef] [Google Scholar]
- Silica nanoparticles promote a-synuclein aggregation and Parkinson's disease pathology. Front Neurosci. 2022;15:807988.
- [CrossRef] [PubMed] [Google Scholar]
- Silica nanoparticles induce neurotoxicity in striatum region by activating oxidative Stress and Apoptosis-related Gene Expression in Adult Male Rats: Possible Involvement of Nrf2/HO-1 and apoptotic pathways. Research Square; 2022
- [CrossRef] [Google Scholar]
- Signal transduction by reactive oxygen species. J Cell Biol. 2011;194:715.
- [CrossRef] [PubMed] [Google Scholar]
- ROS function in redox signaling and oxidative stress. Curr Biol. 2014;24:R453-62.
- [CrossRef] [PubMed] [Google Scholar]
- Molecular dynamic simulations of oxidized skin lipid bilayer and permeability of reactive oxygen species. Sci Rep. 2019;9:110.
- [CrossRef] [PubMed] [Google Scholar]




