
Translate this page into:
5-hydroxytryptamine-evoked contractile activity of large gut in neonatal albino rats
*Corresponding author: Parul Sharma, Department of Physiology, Institute Of Medical Sciences, Banaras Hindu University, Varanasi, Uttar Pradesh, India. drparulsharmajpr@gmail.com
-
Received: ,
Accepted: ,
How to cite this article: Singh S, Mandal MB, Dixit D, Sharma P. 5-hydroxytryptamine-evoked contractile activity of large gut in neonatal albino rats. Indian J Physiol Pharmacol. 2023;67:270-6. doi: 10.25259/IJPP_519_2022
Abstract
Objectives:
Serotonin, also known as 5-hydroxytryptamine (5-HT), is a monoamine neurotransmitter. It is abundantly present in the gut of mammals. The extent to which it contributes to the contraction of the neonatal gut requires further investigation. This study aimed to assess the effect of 5-HT on the contractions of the large intestine in newborn albino rats.
Materials and Methods:
The colon and rectum samples were collected from neonatal and adult albino rats for analysis. Further, in an organ bath, isometric contractions of these isolated gut segments were recorded, in vitro, using a force transducer and a computerised chart recorder, with and without 5-HT in different groups. The 5-HT-induced contractions were also recorded in gut segments pre-treated with various antagonists.
Results:
5-HT (0.01–10 μM) caused a significantly (P < 0.05) greater contractile response (g/g wet tissue) in neonate rats as compared to adults. The response was greater in the rectum as compared to the colon in both neonates and adults. In neonate rats, ondansetron, a 5-HT3 antagonist, could not block the 5-HT-induced large gut contractions, while, in adult rats, it significantly blocked the 5-HT-evoked gut contractility. Methysergide, a 5-HT1/2/5-7 antagonist, blocked the response in both the adult and neonate rectum.
Conclusion:
The 5-HT-evoked response is mediated through 5-HT3 receptor subtypes in adults but not in neonate colon and rectum, indicating possible changes in the distribution of 5-HT receptors in the colon and rectum during development. Furthermore, atropine (a muscarinic cholinergic blocker) and hexamethonium (a ganglion blocker) could not affect the 5-HT-evoked responses in the neonate or adult rats’ colons or rectum. The effect of 5-HT did not appear to involve cholinergic or enteric ganglionic elements.
Keywords
5-Hydroxytryptamine
Colon
Rectum
Gut
Smooth muscle
Contraction

INTRODUCTION
Serotonin, also called 5-hydroxytryptamine (5-HT), plays a significant role in regulating gastrointestinal (GI) motility. 5-HT is present in the GI tract, the central nervous system, and blood platelets.[1] Most of the 5-HT inside the body is in the enterochromaffin cells of the gut.[2] The pathophysiology of functional gut disorders such as irritable bowel syndrome, carcinoid diarrhoea, and emesis induced by chemotherapy is considered to involve various 5-HT receptor subtypes. Several 5-HT receptor subtypes have been implicated in the regulation of intestinal movement.[3] The 5-HT receptor subtypes, such as 5-HT1, 5-HT3, 5-HT4, and 5-HT7, are known to be involved in different types of gut motility disorders.[2,4] Moreover, knowledge of the role of 5-HT in neonatal gut motility is essential, as 5-HT release is known to be involved in neonatal diarrhoea caused by enhanced motility.[5]
In different species and different gut regions, 5-HT has been shown to induce or inhibit gut contractions through several subtypes of 5-HT receptors.[1,6-10] 5-HT3 receptor stimulation induces intestinal contractions in the rat and mice[6,10] but 5-HT2 receptors cause contraction, and 5-HT4 receptors cause relaxation in the rat stomach.[9] In humans, 5-HT4 receptors play a role in the relaxation of the colon.[11,12] Many of these studies were conducted on adult animals only. Thus, whether the various receptor subtypes are present since birth or develop later is not well established. It is known that gut motility functions differ in neonate and adult rats.[13-16] According to a study,[13] congenital pouch colon associated with anorectal malformation does not show spontaneous contractions but retains acetylcholine and histamine-evoked contractions. An in vitro study on rat gut reported that the contractile responses induced by acetylcholine were comparable in adults and neonates, but the blocking effect of atropine in the colon was more prominent in adults than in neonates. Furthermore, this study showed that the histamine’s mechanism of action differs in adults and neonates, as significant enhancement of contractions by pheniramine was observed only in the adult rectum. Hence, this study indicates the existence of different cholinergic and histaminergic activity in adults and neonates, as well as in rectal and colonic tissue.[14] A study conducted on gastric smooth muscles in rabbits reported age-dependent alterations in agonist potency and efficacy. This may partly clarify the alterations in gastric motility during postnatal development.[15] In another study, 5-HT induced concentration-dependent contractions of the ileum in both adult and non-natal rats, but only in neonatal rats; these contractions were weakened by pre-treatment with atropine.[16]
Therefore, recruitment of receptor subtypes may occur during the postnatal period in attaining the functional maturity of adulthood.
In addition, the disorders of motility of the colon and rectum are an important cause of neonatal mortality and morbidity, and several diseases involve the colorectal region in neonates and infants, leading to intestinal motility problems involving 5-HT mechanisms.[5,17] Therefore, considering the importance of 5-HT in large gut motility and the paucity of studies on neonate’s gut, the present study aimed to assess 5-HT-evoked contractile activity in the large gut (colon and rectum) of neonate and adult rats and understand its mechanism of action.
MATERIALS AND METHODS
Adult (4–6 months old) and neonatal (10–16 days old) albino rats of the Charles Foster strain were recruited in this study. The animals were placed in controlled temperature, humidity, and light conditions. A free supply of food and water was provided to them. The experiments were executed following the guiding principles of the institute’s ethical clearance committee (No. Dean/12-13/CAEC/32).
Parts of the large gut (colon and rectum) were dissected out through a vertical abdominal cut immediately after sacrificing the adult rats (through cervical dislocation), and neonatal rats (through decapitation), and the gut segments were cleaned by flushing out the contents. The cleaned gut segments were placed in a Petri dish containing a cold Krebs-Ringer solution. This solution was supplied with 100% oxygen.[13,14]
The cleaned gut segments were cut into longitudinal strips (12–15 mm). The strips were mounted vertically in an organ bath (12 mL) filled with an oxygenated Krebs-Ringer solution (37°C). At one end, the strip was fixed to a glass tube support, and at the other end, it was fixed to a force transducer (MLT 0210, ADI, Sydney, Australia). An initial tension of 0.25–0.5 g was provided.
A specific technique was used to record contractions, as described in the previous studies.[13,14] The mounted gut strips were left for 30 minutes before baseline recordings were taken. Isometric contractions were recorded and amplified using a bridge amplifier, then digitised with the power lab 4/ST system. The recordings were evaluated with Chart-5 software by AD Instruments. Calibration for tension (0–10 g) was done before and after each recording. Baseline recordings were taken for 30 min before exposing gut segments to various serotonin concentrations (0.01–10 μM) to observe dose response. Different antagonists, such as ondansetron, methysergide, atropine sulphate, and hexamethonium, were used to observe their blocking effects on 1 and 3 μM serotonin-induced contractile responses. The Krebs-Ringer solution was prepared with the following standard composition[13,14], and the pH of the solution was 7.4. A stock solution (5 mM) of serotonin was prepared in distilled water. Stock solutions (1 mM) of ondansetron, methysergide, atropine, and hexamethonium were also prepared in distilled water. Serotonin, methysergide, atropine sulphate, and hexamethonium were obtained from Sigma Aldrich Chemicals Pvt. Ltd., India, and ondansetron (Emiset) was attained from Cipla Pharmaceuticals, Mumbai, India. Dilutions of stock solutions were prepared in Krebs-Ringer solution just before the experiments.
Among two sets of experiments, one set of experiments was done to obtain a dose-response of 5-HT. Initially, spontaneously occurring contractions were recorded from the large gut (colon and rectum) strips of adult (n = 6) and neonate (n = 6) rats. Following this, the tissue was treated with each concentration of serotonin at 0.01, 0.1, 1, 3, and 10 μM for 10 min to obtain the dose response. The other set of experiments was performed with various antagonists. Colonic and rectal preparations (n = 6 each for neonates and adults) were exposed to antagonists such as ondansetron, methysergide, atropine, and hexamethonium for 10 min. Subsequently, serotonin (1 or 3 μM) was added for 10 min to see the blocking effect of antagonists.
The recordings of contractions were saved on the personal computer and retrieved later for further analysis. The contractions were expressed as the highest contractile tension (g/g wet tissue) developed after drug treatment.
The mean ± standard error of the mean value of the highest tension (g/g wet tissue) was calculated. The dose-response relationship between the two groups was compared using a two-way Analysis of Variance (ANOVA) (Graph Pad Prism 4 software), and Student’s t -test was performed for paired observations. P < 0.05 was considered significant.
RESULTS
Effect of 5-HT on the contractile activity of large gut (colon and rectum) of neonate and adult rats
Neonatal rat colon 5-HT (0.01–10 μM) produced a dose-dependent enhancement in contractile tension. In the neonatal rectum, 5-HT produced an acute increase in contractile activity with a lower concentration (0.01–1 μM). The response was greater in the rectum in comparison to the colon (P < 0.05, two-way ANOVA). Similarly, in the colon and rectum of adult rats, 5-HT (0.01–10 μM) brought enhancement in contractile activity in a dose-dependent manner with a significantly higher response in the rectum (P < 0.05, two-way ANOVA).
Further, while comparing the 5-HT responses of the neonate with adult rats, it was found that the contractile tension (g/g wet tissue) developed in neonate rats, both colon and rectum, was significantly higher [P < 0.05, two-way ANOVA, Figures 1 and 2].
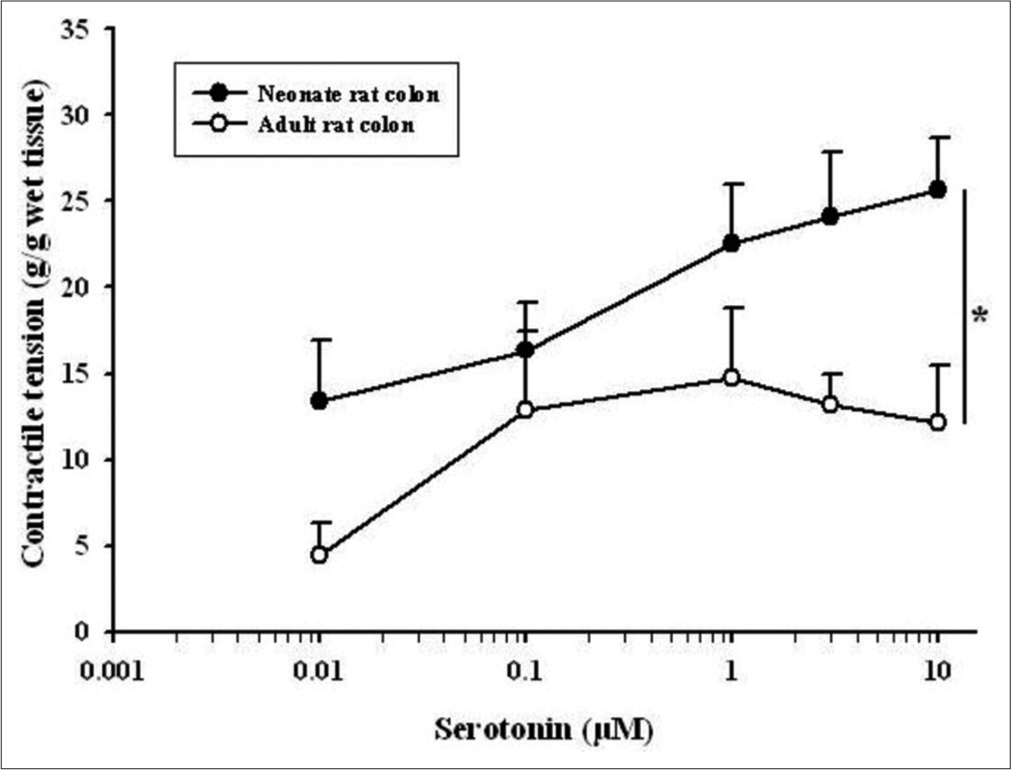
- Dose-response curve showing the comparison of 5-hydroxytryptamine (0.01–10 μM)-induced contractile tension (g/g wet tissue) between the colon of adult and neonate rats. Data points indicate the mean ± standard error of the mean values (n = 6). An asterisk indicates a significant difference (P < 0.05, two-way Analysis of Variance).
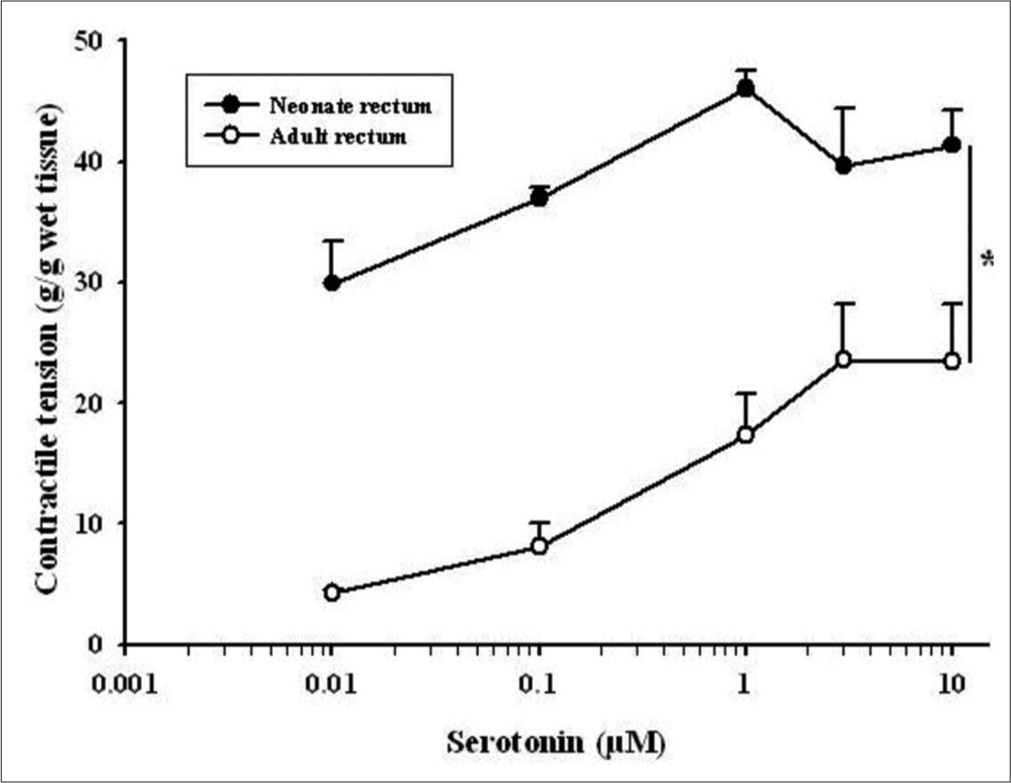
- Dose-response curve showing the comparison of 5-hydroxytryptamine (0.01–10 μM)-induced contractile tension (g/g wet tissue) between the rectum of adult and neonate rats. Data points indicate the mean ± standard error of the mean values (n = 6). An asterisk indicates a significant difference (P < 0.05, twoway Analysis of Variance).
Samples of original recordings of 5-HT (0.01–10 μM)-induced contractions of the colon and rectum of neonate and adult rats are shown in [Figures 3-6].
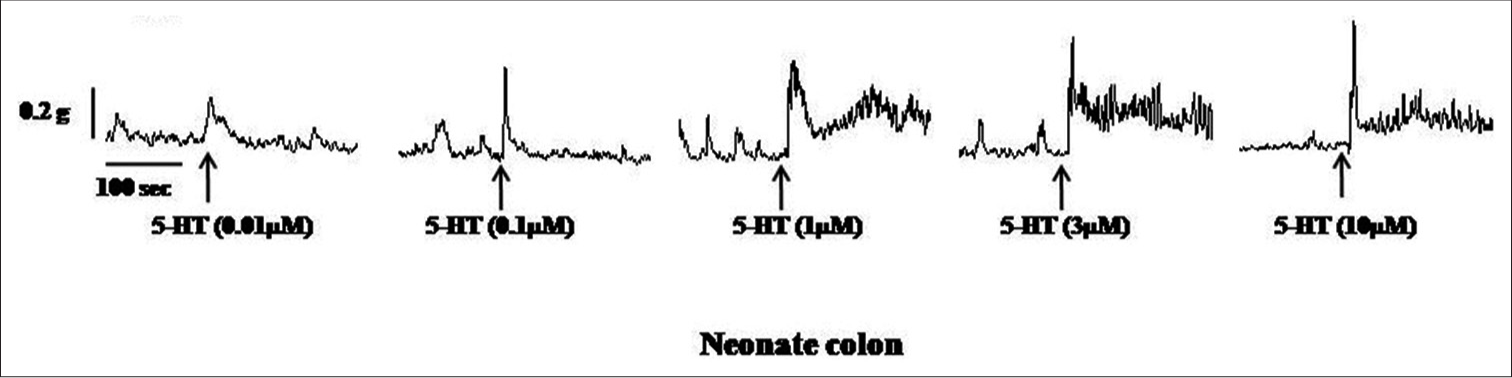
- Samples of the original recording of 5-hydroxytryptamine (5 HT) (0.01–10 μM)-induced contractions of colon a from neonate rats.
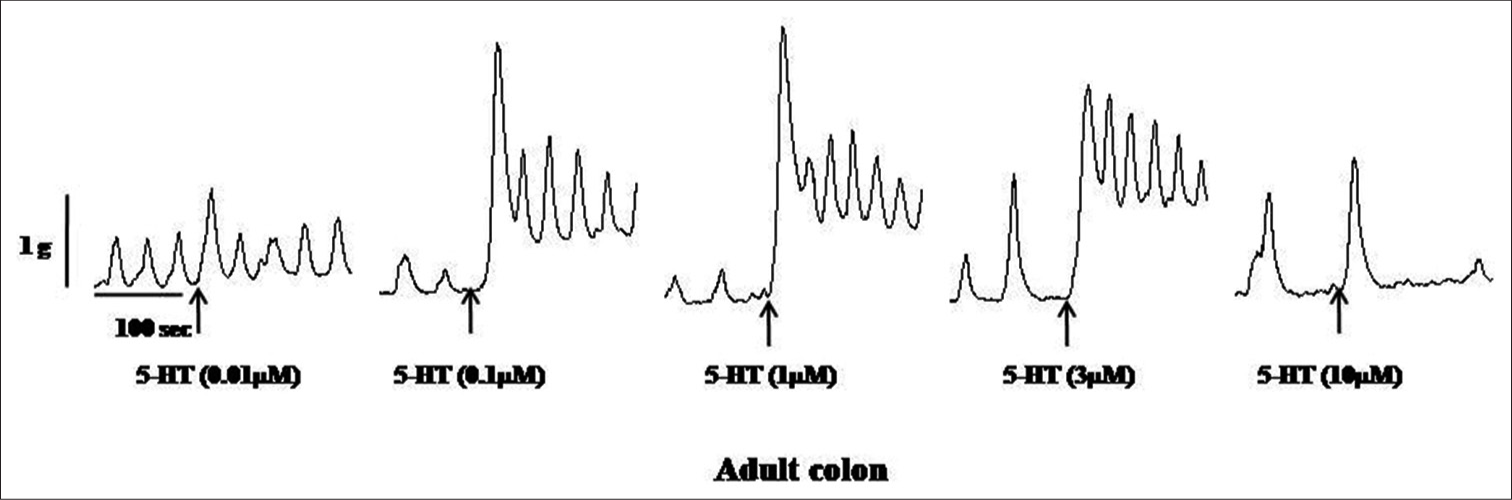
- Samples of the original recording of 5-hydroxytryptamine (5 HT) (0.01–10 μM)-induced contractions of the colon from adult rats.
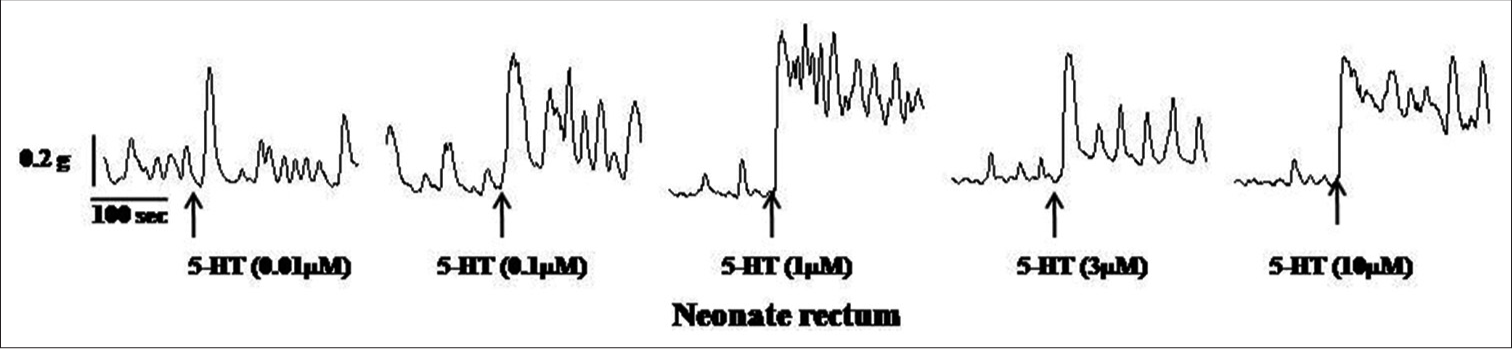
- Samples of the original recording of 5-hydroxytryptamine (5 HT) (0.01–10 μM)-induced contractions of the rectum from neonate rats.
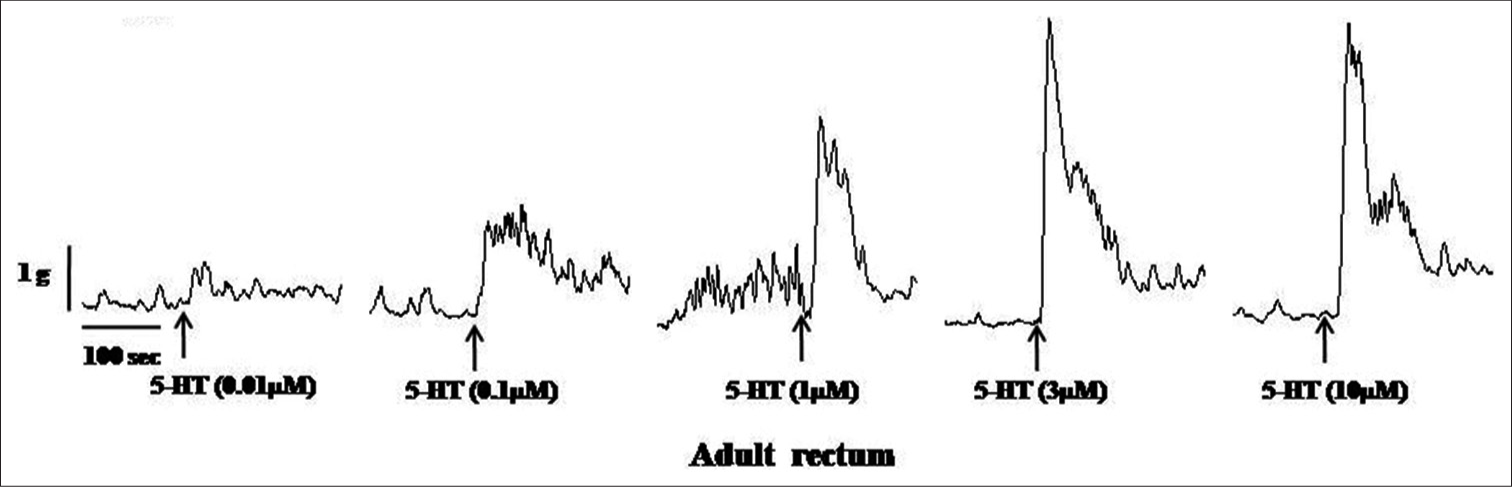
- Samples of the original recording of 5-hydroxytryptamine (5 HT) (0.01–10 μM)-induced contractions of the rectum from adult rats.
The EC50 for 5-HT-induced response in the neonate rat colon was 0.008 μM versus 0.0045 μM in the rectum, indicating that the rectum is two times more sensitive to 5-HT than the colon. In adult rats, EC50 was similar, that is, 0.07 μM both the colon and rectum. The EC50 for 5-HT in both the colon and rectum was nearly ten times greater in neonates as compared to adults.
Effect of antagonists of 5-HT on 5-HT (1 and 3 μM) evoked contractile activity in the large gut (colon and rectum) of adult and neonate rats
In neonate rats, ondansetron (1 μM), a 5-HT3 antagonist, could not block the serotonin (1 and 3 μM) evoked contraction in both the colon and rectum (P > 0.05, Student’s paired t -test). In adult rats, prior administration of ondansetron (1 μM) significantly blocked the 5-HT (1 and 3 μM) evoked contractility in both the colon and rectum (P< 0.05, Student’s paired t -test), [Table 1].
| Antagonists | Before/after antagonists | Contractile tension (g/g wet tissue) for 5-HT (1 µM) *(P<0.05=statistically significant) | Contractile tension (g/g wet tissue) for 5-HT (3µM) *(P<0.05=statistically significant) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Neonate | Adult | Neonate | Adult | ||||||
| Colon | Rectum | Colon | Rectum | Colon | Rectum | Colon | Rectum | ||
| Ondansetron (1 µM) | Before | 8.49±1.86 | 22.54±4.35 | 14.06±2.04 | 14.10±3.84 | 16.94±2.84 | 34.94±4.83 | 14.75±3.19 | 18.92±4.94 |
| After | 7.15±1.26 | 22.91±2.94 | 6.84±2.52* | 5.43±2.38* | 14.99±2.36 | 32.33±3.23 | 8.83±2.02* | 7.86±2.98* | |
| Methysergide (1 µM) | Before | 8.49±1.86 | 22.55±4.35 | 13.60±2.33 | 11.00±3.15 | 17.02±2.81 | 34.42±5.26 | 15.28±3.85 | 18.91±4.94 |
| After | 6.55±1.94 | 7.06±2.79* | 8.56±2.34 | 4.09±1.25* | 13.94±2.79 | 17.73±2.89* | 8.43±2.19* | 10.89±2.43* | |
| Atropine (1 µM) | Before | 28.92±6.48 | 46.68±8.49 | 19.90±6.41 | 18.11±6.09 | 37.89±8.64 | 54.59±10.22 | 26.85±8.32 | 20.71±7.98 |
| After | 27.30±7.89 | 37.61±12.04 | 22.19±9.36 | 12.43±4.13 | 28.83±10.76 | 43.19±12.11 | 25.08±9.53 | 19.93±6.32 | |
| Hexamethonium (1 µM) | Before | 28.92±6.48 | 46.68±8.49 | 19.90±6.41 | 18.11±6.09 | 37.89±8.64 | 54.59±10.22 | 26.85±8.32 | 20.71±7.98 |
| After | 32.38±11.39 | 44.89±14.79 | 21.54±5.73 | 19.62±4.87 | 32.49±11.84 | 51.02±15.51 | 23.89±9.57 | 19.28±6.97 | |
5-HT: 5-hydroxytryptamine, SEM: Standard error of the mean. *P<0.05 (Student's paired t-test, n=4–8)
In neonate rats, methysergide (1 μM), a 5-HT1/2/5-7 antagonist, decreased 5-HT (1 and 3 μM)-evoked contractile tension in the rectum only (P< 0.05, Student’s paired t-test), without any significant change in the colon. On the other hand, in adult rats, methysergide (1 μM) significantly blocked 5-HT (1 and 3 μM)-induced contractions in the rectum (P < 0.05, Student’s paired t -test). Conversely, in adult rat colon, methysergide at a higher concentration (3 μM) significantly inhibited serotonin-induced response (P < 0.05, Student’s paired t-test), [Table 1].
Effect of cholinergic antagonist atropine (1 μM) and ganglionic blocker hexamethonium on 5-HT (1 and 3 μM)-induced contractions
Pre-treatment of the sample with atropine (1 μM) did not alter the 5-HT (1 and 3 μM)-evoked contractions in either the colon or rectum of neonate and adult rats (P > 0.05, Student’s paired t -test), [Table 1].
Similarly, hexamethonium (100 μM) also did not affect the 5-HT (1 and 3 μM)-induced contractions in either the colon or rectum of neonate and adult rats (P > 0.05, Student’s paired t -test), [Table 1].
DISCUSSION
The present investigation demonstrated the effect of 5-HT on the contractile activity of large gut in neonate and adult rats and also explored the possible mechanisms of its action. It was observed that 5-HT caused dose-dependent contractile response in the large gut (rectum and colon) of neonates as well as adult rats at lower doses. Although there was plateau/saturation/decrements in responses with the higher doses, the final dose response (response with the highest dose) was higher than the response with the lowest dose in all the groups. Earlier, 5-HT-induced contractile responses have been examined in small gut, and similar dose-dependent responses in adult and neonate rats were reported.[14-15] Further, in the present investigation, a difference between the contractile response of colon and rectum was explored. It was found that 5-HT caused a higher contractile response in the rectum than in the colon in both adult and neonate rats. This difference in the responses between the colon and rectum may be due to the greater sensitivity of the rectum to 5-HT. Such difference in sensitivity was also reported with capsaicin in the large intestine of a mouse, where the rectum was found to possess a high abundance of capsaicin receptors as compared to other parts of the large gut.[18] A similar phenomenon can be speculated in the present study, too, where the 5-HT receptors population may be higher in the rectum. However, it can be ascertained firmly only after visualising the 5-HT receptor population in the colon and rectum separately.
When the 5-HT-induced contractions were compared between the adult and neonate, a higher contractile response (g/g wet tissue) was recorded in the neonate colon and rectum than in the adult [Figures 1 and 2]. Similarly, in rabbit gastric and colonic smooth muscle, serotonin was found to be more potent in neonates than weanlings.[16,19] Thus, it appeared that neonates’ large gut smooth muscles are more sensitive to serotonin, and during the development process, this sensitivity is blunted.
To assess the receptors involved in 5-HT action, two antagonists of 5-HT (ondansetron and methysergide) were used in the present investigation. In adult rats, pre-treatment with ondansetron, an antagonist of the 5-HT3 receptor, attenuated the contractile response of 5-HT in the colon and rectum. Whereas pre-treatment with methysergide (1 μM), an antagonist of 5-HT1/2/5-7 receptor, inhibited contractile responses evoked by both 1 and 3 μM concentration of 5-HT in the rectum and that by only three μM of 5-HT in the colon of an adult. Adult rat ileum and colon studies revealed a similar response to methysergide and ondansetron.[8,10,16,19,20] However, the effect of these antagonists on the rectum of either adult or neonate was not clear in these reports.
In the present study, ondansetron could not block 5-HT-evoked responses in either the colon or rectum of neonates. In an earlier immunohistochemical study with the small intestine of a 1-day-old rat, 5-HT3 receptor expression was observed substantially in the stomach but not in the intestinal tissue.[20] Therefore, the present observation may be due to less number of 5-HT3 receptors in the neonate gut, and the contractile response is possibly mediated through subtypes of 5-HT receptors other than 5-HT3. Methysergide (1 μM) pre-treatment inhibited 5-HT response in neonate rat rectum only, but not in colon. This indicated that, in neonates, there is an appearance of 5-HT1/2/5-7 in the rectum earlier than the colon. Therefore, expression of 5-HT receptors in the colon may occur late during development, especially in view of the fact that the blocking effect of methysergide was seen in the adult colon.
Further, atropine and hexamethonium were used to assess the involvement of cholinergic or neuronal mechanisms in mediating 5-HT-induced responses. Our results indicated that there may not be any involvement of either cholinergic or enteric neuronal elements because both atropine and hexamethonium failed to block the 5-HT-mediated responses. As observed in the present investigation, the previous studies with adult rat ileum and colon also reported that 5-HT-induced response could not be altered by atropine.[19.20] Thus, the 5-HT-induced contractile responses may be entirely mediated through some other unknown excitatory 5-HT receptor subtypes located in gut smooth muscles. A comparatively recent study has also reported the role of 5-HT3 and 5-HT4 receptors in regulating motor patterns in rat colon.[21]
The exact mechanism behind the greater contractile response of the rectum, as compared to the colon, to 5-HT in both neonate and adult rats needs to be further explored. Furthermore, the mechanism of higher contractile response of neonatal large gut to 5-HT, as compared to adult large gut, requires additional studies. Although we report that in neonatal large gut, 5 HT-induced increase in contractions is not mediated by 5HT3 receptors, more studies are needed to explore the type of receptors through with 5-HT-induced contractile response which is mediated in neonate large gut.
CONCLUSION
In both the colon and rectum of neonate rats, 5-HT produced an increase in contractile response. The response was greater in the rectum as compared to the colon. Similarly, in the adult rat, 5-HT produced an increase in contractile activity in both the colon and rectum, with a significantly higher response in the rectum. Further, while comparing the 5-HT responses of the neonate with adult rats, it was observed that contractile tension (g/g wet tissue) developed significantly more in both the colon and rectum of neonate rats. The response is mediated through 5-HT3 receptor subtypes in adults but not in neonate colon and rectum, indicating possible changes in the distribution of 5-HT receptors in the colon and rectum during development. As both atropine and hexamethonium failed to block the 5-HT-mediated responses, the action of 5-HT seemed to be independent of cholinergic and enteric ganglionic elements. Thus, the 5-HT-induced contractile responses may be entirely mediated through some other unknown excitatory 5-HT receptor subtypes located in gut smooth muscles.
It may, therefore, be concluded that the extent to which 5-HT contributes to the contraction of the large gut is different in neonates and adults. In addition, the mechanism through which 5HT evokes large gut contractions may differ in neonates as it appears to involve different receptors.
Ethical approval
Institutional review board (IRB) permission obtained for the study, No. Dean/12-13/CAEC/32 dated 30.06.2012.
Declaration of patient consent
The authors certify that they have obtained all appropriate consent.
Conflicts of interest
There are no conflicts of interest.
Use of artificial intelligence (AI)-assisted technology for manuscript preparation
The authors confirm that there was no use of artificial intelligence (AI)-assisted technology for assisting in the writing or editing of the manuscript and no images were manipulated using AI.
Financial support and sponsorship
UGC.
References
- Species differences in the 5-hydroxytryptamine-induced contraction in the isolated distal ileum. Jpn J Pharmacol. 1997;74:267-74.
- [CrossRef] [PubMed] [Google Scholar]
- Review article: Serotonin receptors and transporters-roles in normal and abnormal gastrointestinal motility. Aliment Pharmacol Ther. 2004;20(Suppl 7):3-14.
- [CrossRef] [PubMed] [Google Scholar]
- 5-hydroxytryptamine (serotonin) in the gastrointestinal tract. Curr Opin Endocrinol Diabetes Obes. 2013;20:14-21.
- [CrossRef] [PubMed] [Google Scholar]
- Irritable bowel syndrome: New agents targeting serotonin receptor subtypes. Drugs. 2001;61:317-32.
- [CrossRef] [PubMed] [Google Scholar]
- Serotonin and vasoactive intestinal peptide antagonists attenuate rotavirus diarrhea. Gut. 2004;53:952-7.
- [CrossRef] [PubMed] [Google Scholar]
- Activation of 5-HT3 receptors in the rat and mouse intestinal tract: A comparative study. Br J Pharmacol. 2006;148:1012-21.
- [CrossRef] [PubMed] [Google Scholar]
- Characteristics of 5-hydroxytryptamine receptors involved in contraction of feline ileal longitudinal smooth muscle. Korean J Physiol Pharmacol. 2011;15:267-72.
- [CrossRef] [PubMed] [Google Scholar]
- Characterization of the 5-hydroxytryptamine receptors mediating contraction in the intestine of Suncus murinus. Br J Pharmacol. 1999;127:1867-75.
- [CrossRef] [PubMed] [Google Scholar]
- Pharmacological characterization of 5-Hydroxytryptamine-receptor subtypes in circular muscle from the rat stomach. Biol Pharm Bull. 2007;30:508-13.
- [CrossRef] [PubMed] [Google Scholar]
- Pharmacological characterization of the 5-HT receptor-mediated contraction in the mouse isolated ileum. Br J Pharmacol. 2000;131:1716-22.
- [CrossRef] [PubMed] [Google Scholar]
- Characterization of the 5-hydroxytryptamine receptor type involved in inhibition of spontaneous activity of human isolated colonic circular muscle. Br J Pharmacol. 1994;113:143-50.
- [CrossRef] [PubMed] [Google Scholar]
- Differences in response to 5-HT4 receptor agonists and antagonists of the 5-HT4-like receptor in human colonic circular smooth muscle. Br J Pharmacol. 1995;115:172-6.
- [CrossRef] [PubMed] [Google Scholar]
- Pouch colon associated with anorectal malformations fails to show spontaneous contractions but responds to acetylcholine and histamine in vitro. J Pediatr Surg. 2009;44:2156-62.
- [CrossRef] [PubMed] [Google Scholar]
- In vitro study of acetylcholine and histamine induced contractions in colon and rectum of adult and neonate rats. Indian J Physiol Pharmacol. 2013;57:104-13.
- [Google Scholar]
- Developmental changes in agonist-mediated gastric smooth muscle contraction in the rabbit. Pediatr Res. 1989;26:458-61.
- [CrossRef] [PubMed] [Google Scholar]
- The involvement of the serotonergic transmission system in neonatal and adult rat ileum contractility varies with age. Pharmacology. 2011;88:225-32.
- [CrossRef] [PubMed] [Google Scholar]
- Localization of TRPV1 and contractile effect of capsaicin in mouse large intestine: High abundance and sensitivity in rectum and distal colon. Am J Physiol Gastrointest Liver Physiol. 2009;297:348-60.
- [CrossRef] [PubMed] [Google Scholar]
- 5-HT4 receptor agonists: Similar but not the same. Neurogastroenterol Motil. 2008;20:99-112.
- [CrossRef] [PubMed] [Google Scholar]
- Contraction of the rat colon by sympathomimetic amines: Effect of methysergide and 5-HT desensitization. Eur J Pharmacol. 1972;17:319-24.
- [CrossRef] [PubMed] [Google Scholar]
- Involvement of 5-HT3 and 5-HT4 receptors in colonic motor patterns in rats. Neurogastroenterol Motil. 2015;27:914-28.
- [CrossRef] [PubMed] [Google Scholar]




